
天门冬酰胺酶基因在黑曲霉中的同源表达
2015-01-06 09:45:20韦素真岳苗苗
食品与生物技术学报 2015年5期
韦素真, 张 会, 姚 杨,岳苗苗,李 杰
(东北农业大学 生命科学学院,黑龙江 哈尔滨 150030)
天门冬酰胺酶基因在黑曲霉中的同源表达
韦素真, 张 会, 姚 杨,岳苗苗,李 杰*
(东北农业大学 生命科学学院,黑龙江 哈尔滨 150030)
根据NCBI中的黑曲霉天门冬酰胺酶基因asp序列(GI:145231287)设计特异性引物,从黑曲霉CICC2462基因组中扩增asp基因编码区,构建其黑曲霉表达载体pSZHG-Asp。通过农杆菌介导法转化黑曲霉,筛选以asp基因置换糖化酶基因glaA的同源重组转化子。对转化子的表达产物进行SDS-PAGE分析和酶活检测。获得8株在glaA位点发生基因置换的同源重组转化子,并对其中4株的上清液进行检测。SDS-PAGE结果显示,在42 000处有目的蛋白质条带,重组菌株的目的蛋白质表达量为185~417 μg/mL,发酵液最高酶活为21 111 U/mL。
天门冬酰胺酶;黑曲霉;基因置换;Berthelot显色反应
天门冬酰胺酶(Asparaginase)又叫天冬酰胺酶、 L-天门冬酰胺酶、门冬酰胺酶等,它是一种催化天门冬酰胺水解生成天门冬氨酸和氨的酶,广泛存在于植物、动物组织及微生物中[1]。来自于黑曲霉的天门冬酰胺酶是一种食品添加剂,其功能是降低油炸或膨化食品中的丙烯酰胺含量。2002年4月,瑞典国家食品管理局和斯德哥尔摩大学研究人员率先报道,在一些烧烤和油炸的淀粉类食品中,例如油炸土豆片、薯条等中检出丙烯酰胺,而且含量超过饮水中允许最大限量的500多倍[2]。之后英国、瑞士、挪威和美国等国家也相继报道了类似结果[3]。丙烯酰胺具有遗传毒性、神经毒性和潜在致癌性,因此控制食品中丙烯酰胺含量尤其重要[4]。丙烯酰胺主要是通过美拉德反应,由食品原料中的天冬酰胺和还原糖在高温加热过程中形成的[5],天门冬酰胺酶可以将丙烯酰胺的前体物——天冬酰胺转化成天冬氨酸,从而减少丙烯酰胺的含量[6],可以从源头上控制丙烯酰胺的生成[7]。荷兰的帝斯曼公司成功地研发了一种酶制剂——PreventASeTM,经过天冬酰胺酶反应过程,可有效消除90%以上的丙烯酰胺成分,并申报了专利[8]。2005年我国卫生部发布公告,要求人们注意食品中的丙烯酰胺的含量,呼吁采取积极的措施减少其含量[9],并在2009年把天门冬酰胺酶列为食品添加剂[10],但是国内关于天门冬酰胺酶作为食品添加剂的研究和应用报道很少。作者拟构建食品级天门冬酰胺酶工程菌,为天门冬酰胺酶的工业化生产和应用奠定基础。
1 材料与方法
1.1 材料
1.1.1 菌株与载体黑曲霉CICC2462:由肇东市日成酶制剂有限公司提供;农杆菌AGL1:由作者所在实验室保存;大肠杆菌菌株E.coli DH5α:购自北京全式金生物技术公司。载体pSZHG:由作者所在实验室保存;载体pSZHG-Asp:由作者所在实验室构建;PMD18-T vector:购自大连宝(TaKaRa)生物工程公司。
1.1.2 试剂各种限制性内切酶、T4DNA连接酶、Taq酶、核酸相对分子质量标准、质粒提取试剂盒、DNA回收试剂盒等:购自TaKaRa公司;蛋白质相对分子质量标准、琼脂糖等:购自BBI公司;氨苄青霉素、卡纳抗生素、潮霉素B、蛋白胨、氯化钙、氯化钠、柠檬酸钠、水杨酸、亚硝基铁氰化钠、天门冬酰胺等:均为国产试剂。
1.1.3 培养基LB培养基、YEB培养基、PDA培养基、发酵培养基:均按照常规实验室标准配置[11]。
1.2 方法
1.2.1 黑曲霉基因组DNA提取黑曲霉基因组的提取方法采用CTAB方法[12]。
1.2.2 引物设计根据Genebank中公布的黑曲霉天门冬酰胺酶基因Asp序列 (序列号GI:145231287)以及黑曲霉表达载体pSZHG多克隆位点的特征,用Primer5.0软件分别设计Asp基因特异引物。根据黑曲霉表达载体pSZHG-Asp的序列以及黑曲霉糖化酶基因位点的序列,设计转化子、PCR检测引物及同源重组转化子PCR检测引物。引物序列见表1。
1.2.3 PCR反应PCR基本的反应体系如下(25 μL):5×Buffer 5 μL,Mg2+(25 mmol/L)2 μL,dNTP(2.5 mmol/L)2 μL,上下游引物(20 μmol/L)各0.5 μL,模板 1 μL,Prime Star Taq酶 0.2 μL,加ddH2O至25 μL。
PCR反应条件:94℃预变性5 min,94℃变性30 s,56℃退火30 s,72℃延伸2 min,共30个循环,72℃终延伸10 min,4℃终止反应。
1.2.4 表达载体构建将pSZHG、pMD-Asp载体分别用NheⅠ、HindIII限制性内切酶酶切,分别回收15 000 bp的载体条带和1 254 bp的目的基因条带,连接并构建表达载体pSZHG-Asp。

表1 本研究所用引物Table 1 Primers used in this research
1.2.5 黑曲霉的遗传转化见文献[12]。
1.2.6 采用根癌农杆菌介导法见文献[13]。
1.2.7 天门冬酰胺酶酶活测定活性测定以及相关试剂的配置参照文献[14-15]。标准曲线绘制:精确称取(NH4)2SO4溶于超纯水,配置质量浓度分别为5、10、20、50、70、100 μg/mL的标准溶液。按表2进行标准曲线的制作以及天门冬酰胺酶活性的测定[16]。

表2 标准曲线制作Table 2 Standard curve
在各管中加入1.2 mL水杨酸溶液,再加入0.10 mL亚硝基铁氰化钠溶液和0.10 mL次氯酸钠溶液,混匀,37℃水浴30 min。用1 cm比色皿,按照表3标准管列分别加入不同浓度的标准溶液,反应后测定650 nm波长的吸光度,以水作参比,测定各管溶液的吸光度。

表3 天门冬酰胺酶酶活的测定步骤Table 3 Procedure for asparaginase determination
混匀后置37℃水浴30 min,于650 nm处用无氨水调零,测定各管吸光度。测定管吸光度减去对照管吸光度所得的吸光度值,对照标准曲线得到脱氨反应产生的氨质量浓度X(μg/mL)计算酶活[17]:
酶活 (μg/(min·mL))=(X μg/30 min)×(1/0.03 mL)=10X/9
酶活性单位定义(U):在标准条件下,1 mL酶液在37℃与底物作用1 min产生1 μg氨为1个酶活性单位(pH 5.0,37℃)。
2 结果与讨论
2.1 asp基因的扩增
以黑曲霉CICC2462基因组为模板,用asp基因特异性引物进行PCR扩增,获得约1 254 bp的目的片段,见图1。将该片段克隆至pMD18-T,送交测序。将测序结果进行blast比对分析发现,该序列与登录号 GI:145231287的基因序列相似性为100%。

图1 asp基因PCR扩增结果Fig.1 Results of PCR amplification asp
2.2 表达载体构建
表达载体pSZHG-asp构建见图2。该载体用XbaI、HindIII双酶切鉴定,得到大约14 000 bp和2 300 bp的两条带,见图3,证明构建正确。

图2 pSZHG-asp表达载体的构建Fig.2 Construction of expression vector pSZHG-asp

图3 pSZHG-asp双酶切结果Fig.3 Enzymes digestion of vector pSZHG-asp
2.3 同源重组转化子的筛选
农杆菌与黑曲霉共培养之后,挑取筛选平板长出的黑曲霉菌落,摇菌,提取基因组。利用P3、P4引物进行PCR检测,共对19个菌株进行检测,有4株扩增出3 500 bp左右的目的条带和4 700 bp的glaA基因条带,为阳性转化子,见图4。利用P5、P6引物对4株阳性转化子做PCR检测,结果均能扩增出1 700 bp左右的目的条带,因此这4株都是同源重组转化子,见图5。
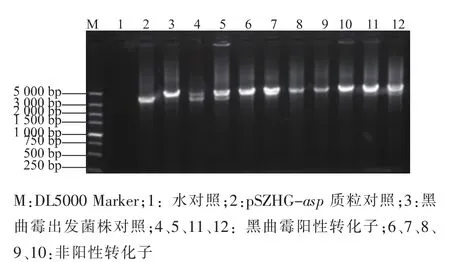
图4 黑曲霉转化子PCR验证结果Fig.4 Results of PCR of Aspergillus niger transformants
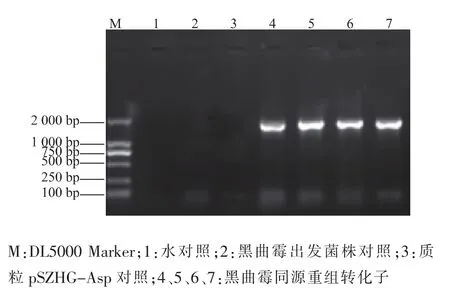
图5 黑曲霉转化子同源重组PCR验证结果Fig.5 Results of PCR of homologous recombination Aspergillus niger transformants
2.4 天门冬酰胺酶酶活测定
2.4.1 氨态氮标准曲线的制作按表1标准管系列分别加入不同质量浓度的标准溶液,反应后在650 nm处测定吸光度,以氨质量浓度(μg/mL)为横坐标,吸光度为纵坐标,绘制标准曲线,见图6。R2= 0.999 7,表明其线性相关性较好。
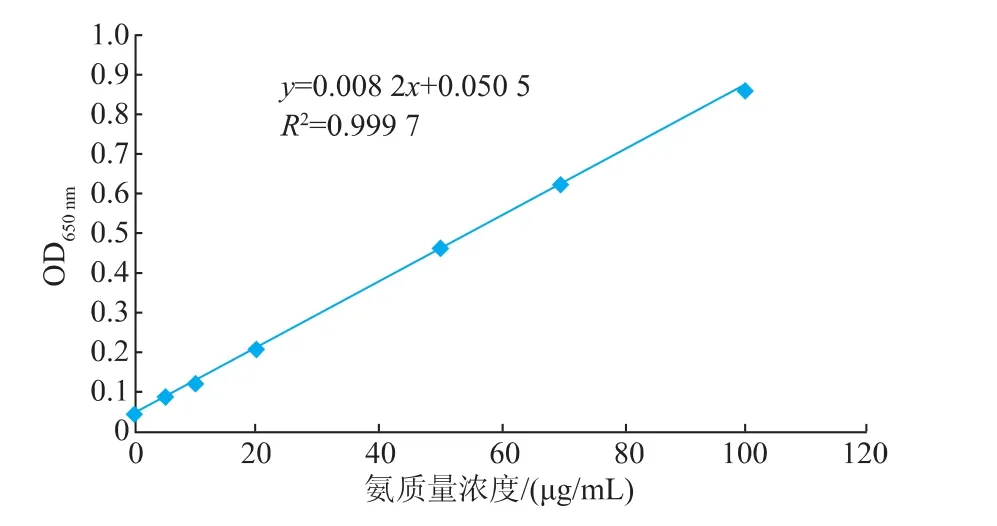
图6 氨态氮标准曲线Fig.6 Standard curve of ammonium nitrogen assay
2.4.2 酶活测定取出发菌株和3个同源重组菌株进行摇瓶发酵培养,不同发酵时间的酶活见表4。最高酶活可达21 111 U/mL。
2.5 天门冬酰胺酶在重组菌株中的分泌表达
对培养发酵10、11 d的菌液,取上清液进行SDS-PAGE分析,结果见图7。与出发菌株相比,3株重组菌株都在约42 000的位置出现了目的蛋白质条带。经ALPHAIMAGER SYSTEMS软件分析,3株重组菌株的目的蛋白质表达量为 185~417 μg/mL,而出发菌株仅为12 μg/mL。
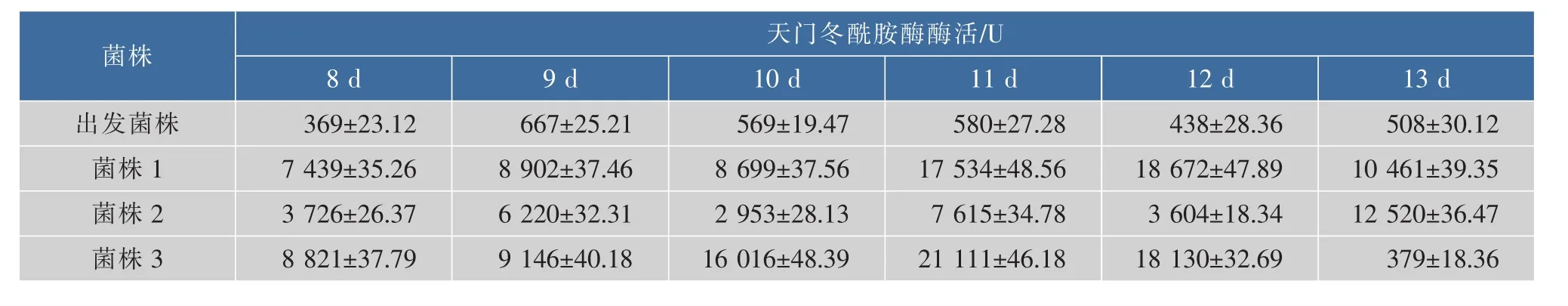
表4 天门冬酰胺酶在不同发酵时间的酶活Table 4 Enzyme activity detection at different fermentation time of asparaginase
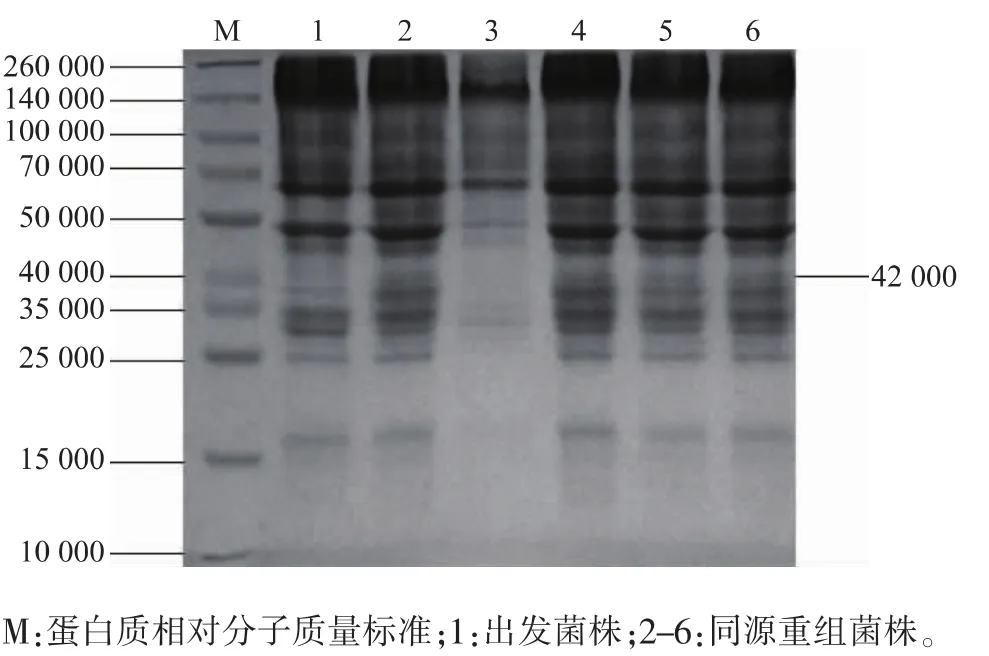
图7 表达蛋白SDS-PAGEFig.7 SDS-PAGE of expressed protein
3 结语
随着生活质量的提高,食品安全问题越来越引起人们的关注。丙烯酰胺具有遗传毒性、神经毒性和潜在致癌性。我国已将食品中丙烯酰胺的检验列为“十五”国家重大科技专项“食品安全关键技术”课题。降低食品中丙烯酰胺的方法有很多,但是使用天门冬酰胺酶降解原料中的天门冬酰胺是最为简便有效的方法。因此,天门冬酰胺酶已在2009年被国家卫生部确定为一种食品添加剂。但是我国目前还没有制定各种食品中丙烯酰胺的限量标准,天门冬酰胺酶的研究和应用还没有得到足够的重视。
作者以糖化酶生产菌株CICC2462同时作为基因供体和转化受体,以糖化酶基因位点为基因整合的靶位点,构建了天门冬酰胺酶工程菌,且在筛选标记潮霉素基因表达框两侧设计了顺向重复序列,可在后续实验中删除筛选标记基因,达到食品级表达系统要求。本研究获得的工程菌摇瓶发酵酶活最高已达到20 000 U/mL以上,具有明确的产业化的前景。但是目的蛋白质产量只有417 μg/mL,这说明表达的天门冬酰胺酶具有较高的比活,同时也有巨大的提升表达量的潜力。
[1]张卓丹,赵蓉,吴涛.控制食品中丙烯酰胺含量的方法研究进展[J].食品科学,2009,30(1):294-297.
ZHANG Zhuodan,ZHAO Rong,WU Tao.Summary of control metheds for reducing acrylamide in food[J].Food Science,2009,30(1):294-297.(in Chinese)
[2]Tareke E,Rydberg P,Karlsson S,et al.Acrylamide:cooking carcinogent[J].Chen Res Toxicol,2002(13):517-522.
[3]徐东路,宋贤良.食品中的丙烯酰胺及生物解决方案[J].食品与发酵工业,2010,12:152-155.
XU Donglu,SONG Xianliang.Acrylamide content in food and enzymtic solution[J].Food and Fermentation Industries,2010,12:152-155.(in Chinese)
[4]王文艳,刘凌,周明.天冬酰胺酶降低膨化食品丙烯酰胺含量的初步研究[J].中国食品添加剂,2011,5:59-63.
WANG Wenyan,LIU Ling,ZHOU Ming.Preliminary study on reducing acrylamide content in puffed foods by adding asparaginase[J].China Food Additives,2011,5:59-63.(in Chinese)
[5]Mottram D S,Wedzicha B L,Dodson A T.Acrylamide is formed in the miallard reaction[J].Nature,2002,419:448-449.
[6]杨波.膨化食品及其丙烯酰胺的研究进展[J].食品科技,2011,3:42-45.
YANG Bo.Research progress in studies on puffing food and its acrylamide[J].Food Science&Technology,2011,3:42-45.(in Chinese)
[7]欧什益,张玉萍,黄才欢,等.几种添加剂对油炸薯片中丙烯酚胺产生的抑制作用[J].食品科学,2006,27(5):137-189.
Ou Shiyi,ZHANG Yuping,HUANG Caihuan,et al.Inhibition of acrylamide formation in fried potato crisps by some food additives[J].Food Science,2006,27(5):137-189.(in Chinese)
[8]Monica Anese,Michele Suman M,Cristina Nicoli.Technological strategies to reduce acrylamide levels in heated foods[J].Food Eng Rev,2009,1:169-179.
[9]Hendriksen H V,Kornbrust B A,Oestergaard P R,et al.Evaluating the potential for enzymatic acrylamide mitigation in a range of food products using an asparaginase from Aspergillus oryzae[J].Journal of Agricultural and Food Chemistry,2009,57(10):4168-4176.
[10]余以刚,李理,梅艳群,等.减少高温加工食品中丙烯酰胺含量的几种方法[J].现代食品科技,2007(1):83-85.
YU Yigang,LI Li,MEI Yanqun,et al.Methods for reducing acrylamide content in high-temperature processed foods[J].Modern Food Science and Technology,2007(1):83-85.(in Chinese)
[11]龙小涛,何嘉锐,叶雪丽.食品中丙烯酰胺的抑制方法研究进展[J].现代食品科技,2012,6:688-691.
LONG Xiaotao,HE Jiarui,YE Xueli.Study on inhibition methods of acrylamide in food[J].Modern Food Science and Technology,2012,6:688-691.(in Chinese)
[12]张谷楠.黑曲霉葡萄糖氧化酶基因在毕赤酵母和黑曲霉中的分泌表达[D].哈尔滨:东北农业大学,2012.
[13]张会.β-1,4-内切木聚糖酶基因在黑曲霉中安全高效表达的研究[D].哈尔滨:东北农业大学,2013.
[14]季艳伟,许杨,黄志兵,等.催化光度法测定三聚氰胺脱氨酶活性[J].食品科学,2011,32(18):234-238.
JI Yanwei,XU Yang,HUANG Zhibing,et al.Determination of melamine deaminase activity by catalytic spectrophotometry[J]. Food Science,2011,32(18):234-238.(in Chinese)
[15]GB/T 18204.25-2000,公共场所空气中氨测定方法[S].
[16]涂追,许杨,季艳伟,等.三聚氰胺脱氨酶基因克隆表达及活性测定[J].食品与技术学报,2011,30(2):234-238.
TU Zhui,XU Yang,JI Yanwei,et al.Cloning,expression and characterization of recombinant melamine deaminase in E.coli[J]. Journal of Food Science and Biotechnology,2011,30(2):234-238.(in Chinese)
[17]何战胜,邓健,许金生,等.催化光度法测定体液腺苷脱氨酶的活性[J].中国卫生检验杂志,2005,15(5):546-547.
HE Zhansheng,DENG Jian,XU Jinsheng,et al.Determination of adenosine deaminase activity in body fluids by catalytic spectrophotometry[J].Chinese Journal of Health Laboratory Technology,2005,15(5):546-547.(in Chinese)
Homologous Expression of Asparaginase Gene in Aspergillus niger
WEI Suzhen, ZHANG Hui, YAO Yang, YUE Miaomiao, LI Jie*
(College of Life Sciences,Northeast Agricultural University,Harbin 150030,China)
In order to achieve the homologous expression of asparaginase gene asp in Aspergillus niger CICC2462,the asparaginase engineering strains were construced.The specific primers were designed according to asparaginase gene asp sequence in the NCBI(GI:145231287)to amplify the coding region of asp and pSZHG-asp expression vector was constructed.Then agrobacterium mediated method was utilized to transform Aspergillus niger,and homologous recombinations which glucoamylase gene(glaA)was replaced by Asp were screened.The expression products of the transformant were analyzed using SDS-PAGE and enzyme activity detection.Eight homologous recombination strains,where glaA locus was replaced,were obtained,and the supernatant in 4 of them were measured.The result of SDS-PAGE showed protein bands at 42 000,the amount of the recombinated protein was 185~417 μg/mL,and the highest enzyme activity of fermentation broth was up to 21 111 U/mL.
asparaginase,Aspergillus niger,gene replacement,berthelot colour reaction
Q 786
A
1673—1689(2015)05—0554—06
2014-02-19
农业生物功能基因重点实验室开放课题资助项目。
*通信作者:李 杰(1972—),男,黑龙江拜泉人,农学博士,副教授,硕士研究生导师,主要从事分子遗传学与基因工程方面的研究。
E-mail:lijie_neau@126.com
猜你喜欢
科学导报(2024年19期)2024-04-22 05:53:32
知识窗(2022年7期)2022-08-05 02:12:35
云南化工(2021年11期)2022-01-12 06:06:18
农民致富之友(2019年12期)2019-05-22 09:30:42
食品与发酵工业(2019年1期)2019-01-29 10:25:08
现代食品(2016年24期)2016-04-28 08:12:08
当代化工研究(2016年2期)2016-03-20 16:21:20
中国酿造(2016年12期)2016-03-01 03:08:22
中国酿造(2016年12期)2016-03-01 03:08:20
大连工业大学学报(2015年4期)2015-12-11 04:06:50
