
嗜线虫致病杆菌HB310 几丁质酶基因chi70 的克隆和表达
2014-12-03 05:55:22王永娟南宫自艳白向宾王勤英
环境昆虫学报 2014年6期
王永娟,张 杰,南宫自艳,宋 萍,白向宾,王勤英
(河北农业大学植物保护学院,保定 071000)
几丁质酶是一类可以降解几丁质的糖苷酶,存在于细菌、真菌、昆虫、甲壳纲动物以及高等动植物体内。在农业生产中,几丁质酶可以降解昆虫体内围食膜中的几丁质,破坏昆虫保护屏障,从而为其他昆虫病原生物侵入到昆虫体内提供便利,提高昆虫感染率和死亡率,研究表明,几丁质酶可以作为增效因子增强Bt 的毒力(Arora et al.2003)。几丁质酶也可抑制真菌孢子萌发和菌丝生长,从链霉菌中分离到的几丁质酶能抑制曲霉菌丝的生长(Frank et al.,2005);从荧光假单胞菌中分离的几丁质酶对棉花枯萎病菌有抑制作用(Naosekpam et al.,2006)。Chen 等从昆虫病原线虫共生菌Xenorhabdus 和Photorhabdus 分离出高活性的几丁质酶,并对其进行酶活和抑菌实验研究(Chen et al.,1996),认为几丁质酶普遍存在于昆虫病原线虫共生菌内,但有关共生菌几丁质酶的作用机制研究报道却并不多。
嗜线虫致病杆菌X.nematophila HB310(Xn HB310)菌株是本实验室从小卷蛾斯氏线虫Steinernema carpocaposae HB310 中分离到的共生菌,对多种包括鳞翅目、鞘翅目在内的农林害虫表现出很强的杀虫活性(李秀花等,2003),具有巨大的开发应用前景。王勤英等从Xn HB310 分离得到多种具有血腔注射活性和胃毒活性的杀虫蛋白,并对这些蛋白的杀虫机理进行了研究(王勤英等,2005;Wang et al.,2012)。本研究通过克隆和表达嗜线虫致病杆菌Xn HB310 几丁质酶基因chi70,对蛋白Chi70 进行初步研究,为进一步研究Xn HB310 杀虫机制和扩大应用范围提供理论依据。
1 材料与方法
1.1 菌株、质粒和试虫
嗜线虫致病杆菌Xn HB310、pET-28a 为本实验室保存,菌株Escherichia coli DH5α 和E.coli Transetta(DE3)购自于北京全式金公司,pMD18-T 购自于TAKARA 公司。棉铃虫Helicoverpa armigera 取自河北农业大学生物防治实验室养虫室。
1.2 主要试剂
PCR 相关试剂、Marker 等购自于北京全式金公司,限制性核酸内切酶购自于TAKARA 公司,DNA 胶回收试剂盒购自于上海生工。
1.3 培养基
LB 培养基:牛肉蛋白胨10 g,酵母浸粉5 g,NaCl 10 g,定容1 L,pH7.2-7.4。
NBTA 培养基:营养琼脂45 g,氯化三苯基四氮唑(TTC)0.025 g,溴百里酚蓝(BTB)0.04 g,定容到1 L,pH7.2-7.4。
牛肉汤培养基:牛肉蛋白胨10 g,牛肉膏3 g,NaCl 5 g,定容1 L,pH7.2-7.4。
1.4 胶体几丁质
2 g 粉末几丁质加入40 mL 浓盐酸,搅拌至完全溶解,800 mL 50%乙醇沉淀过夜,离心,用大量的去离子水洗涤致中性,最后定容到200 mL,制成1%胶体几丁质。
1.5 细菌总DNA 的提取
将保存于NBTA 平板的Xn HB310 菌落接入到牛肉汤培养基中,180 rpm、28℃震荡培养24 h,其菌液用于提取细胞总DNA,提取方法参照(Sanbrook et al.,2003)。
1.6 几丁质酶基因chi70 克隆及原核表达体系的构建
根据NCBI 已知序列及功能预测分析设计基因chi70 克隆引物,上游引物 C70F:5'-ATGTCTCAAAATGTTTATCGATAC-3',下游引物C70R:5'-CTACGATTTACGACGGGTTAC-3'。进行PCR 扩增,条件为:94℃预变性10 min;94℃30 s,56℃40 s,72℃1 min,30个循环;72℃延伸10 min。PCR 产物与pMD18-T 载体连接后,转入E.coli DH5α。PCR 检测正确后进行测序。根据测序结果,重新设计表达引物进行PCR,上游引物chi70F:5'-CGCGGATCCATGTCTCAAAATGT TTATCGATA-3'(BamHⅠ酶切位点),下游引物chi70R:5'-CCCAAGCTTCTACGATTTACGACG GGTTAC(Hind Ⅲ酶切位点)。PCR 产物经BamHⅠ和Hind Ⅲ酶切处理后插入到相同酶切处理的pET-28a 载体,转入到DH5α,构建出表达载体pET-28a-chi70。通过测序对插入的chi70基因进行检测。
1.7 几丁质酶基因chi70 的分子生物信息学分析
利用 ExPASy Translate tool 在线程序和DNAMAN 等软件对该基因进行分子生物学分析。
1.8 几丁质酶的原核表达及诱导条件优化
将构建好的表达载体pET-28a-chi70 转入到E.coli Transetta(DE3)中,转化子经PCR 验证后接入到5 mL 含有50 μg/mL 卡那霉素和氯霉素的LB 培养基中,37℃、180 rpm 培养过夜,次日按照1%接种量接入到200 mL 含有相同抗生素的LB培养基中,37℃、180 rpm 培养至OD600为0.5,加入100 mmol/L IPTG 继续在37℃震荡培养。通过设置不同诱导温度对菌株进行诱导条件的优化。以相同处理的空载体转化子E.coli Transetta(pET-28a)为对照。将诱导的菌液离心,10 mL PBS 悬浮,超声破碎(工作2 s,间隔5 s,共15 min),4℃、10000 rpm 离心15 min,上清经0.22 μm 细菌过滤器得到诱导表达的粗蛋白液,SDS-PAGE 电泳检测,蛋白含量的测定采用考马斯亮蓝法(郭尧君,2001)。
1.9 几丁质酶活性测定
采用二硝基水杨酸法(DNS 法)检测几丁质酶活性(李华等,2003)。设置不同pH 值(4.0、5.0、6.0、7.0、8.0、9.0 和10.0)的缓冲液,求出反应体系中影响几丁质酶活力的最适pH 值。
1.10 几丁质酶增效作用的测定
将Bt HD-73 菌株悬液浓度设定为400、200、20、10、2 和1 μg/mL,分别与等体积的无菌水、含有0.3 g/mL 几丁质酶粗提液加入到24 孔板中,每孔分别接入1 头棉铃虫2 龄幼虫。CK 组分别为灭菌水、Chi70 诱导菌液和Chi70 未诱导菌液的粗提液。每个处理48 头试虫,试虫在26±1℃、相对湿度60%、光周期L∶D 为14∶10 条件下饲养72 h,计算LC50。计算方法参照张志祥(张志祥等,2002)。
2 结果与分析
2.1 几丁质酶基因的克隆和生物信息学分析
以chi70 全长引物从嗜线虫致病杆菌Xn HB310 总DNA 中PCR 扩增出了约1900 bp 条带,测序结果表明chi70 全长基因为含有1947bp 的完整开放阅读框,编码648个氨基酸(见图1),推导的Chi70 蛋白理论分子量为72.4 kDa,等电点4.89,为酸性蛋白,GC 含量48.8%。将该基因序列提交GenBank 登记,登录号为KC701471。
利用ExPASy ProtParam 在线工具对Chi70 蛋白进行分析,结果显示,Chi70 由10017个原子组成,分子式为C3231H4903N869O998S16。Chi70 水溶液在280 nm 处的消光系数为118150,不稳定指数为38.06,半寿期大于10 h(参照大肠杆菌体内值),由此可认为该蛋白在细菌胞内可稳定存在。Chi70的脂溶指数(aliphatic index)为71.10,疏水性平均值(GRAVY)为-0.553。利用Protscale 程序对Chi70 蛋白进行疏水性图谱分析,其大部分氨基酸属于亲水性,由此表明该蛋白为水溶性蛋白。利用SignalP4.0 对Chi70 蛋白序列信号肽的预测结果显示,Chi70 不含信号肽。
将Chi70 氨基酸序列与GenBank 中登录的序列进行比对,从中选择3个共生菌不同菌株几丁质酶氨基酸序列进行同源性比较,结果见图2。从图中可以看出,Xn HB310 的Chi70 蛋白与不同属的共生菌P.asymbiotica 几丁质酶同源性并不高,这可能是由于它们的起源不同,但进化过程属于趋同进化(Chaston et al.,2013);与同种的X.nematophila ATCC19061 的同源性很高;而同属不同种的X.bovienii 的序列差异性也很高,说明几丁质酶在进化上并不保守,正是因为这种变异,共生菌不同菌株在降解几丁质活性上表现出明显差异。
2.2 几丁质酶的原核表达及重组蛋白SDS-PAGE分析
将重组表达载体pET-28a-chi70 转入到Transetta(DE3)后,加入终浓度为1 μmol/L 的IPTG 在37℃进行诱导表达,SDS-PAGE 电泳检测。结果表明,未诱导的菌株没有表达目的蛋白,而诱导的菌株随着诱导时间的延长,目的蛋白表达量增加,在80 kD 左右处可以看见明显的诱导蛋白(见图3)。Chi70 几丁质酶蛋白理论值为72.4 kDa,带有两个HIS 标签共6 kDa,共78 kDa,说明表达菌株正确地表达了目的蛋白。目的蛋白以包涵体的形式存在,通过改变诱导温度使之大量表达出可溶性目的蛋白,当诱导温度为28℃时,大量诱导出可溶性融合蛋白。
2.3 几丁质酶酶活测定
以NAG 为反应底物,设置不同梯度浓度,测定OD540值,以确定NAG 浓度与吸光度之间的标准曲线。得到标准曲线公式为Y=9.8313X-0.0344(R2=0.9905)(其中Y 代表吸光度,X 代表底物浓度)。pH 值对Chi 酶活的影响见图4,可以看出,重组Chi70 几丁质酶活性在pH 值为7.0 时最高,为8.815 U/mL,表明该几丁质酶在中性条件下具有相对较高的酶解活性。
2.4 几丁质酶的增效作用
分别测定Bt HD-73 菌悬液和HD-73+Chi70混合物对棉铃虫二龄幼虫的LC50。结果见表1。从表1 中可以看出,单独用HD-73 饲喂棉铃虫时的致死中浓度LC50为14.80 μg/mL;与Chi70 几丁质酶混合后饲喂棉铃虫幼虫时的致死中浓度LC50则为5.66 μg/mL,后者显著低于前者,即重组几丁质酶Chi70 对Bt HD-73 具有增效作用。

图1 chi70 基因推导的氨基酸序列Fig.1 Deduced amino acid sequence of gene chi70

图2 Chi70 蛋白氨基酸与其他3种几丁质酶序列同源比对Fig.2 Alignment of the amino acid sequence of Chi70 with those of three other homologous species

图3 chi70 基因表达产物SDS-PAGE 分析Fig.3 SDS-PAGE analysis of expressed product of chi70 gene
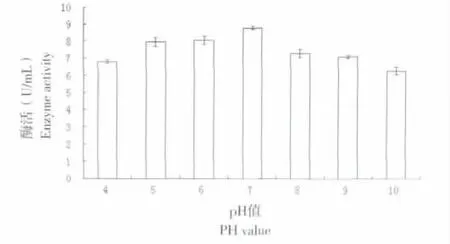
图4 Chi70 几丁质酶在不同pH 值条件下酶活Fig.4 Chi70 chitinase enzyme activity at different pH value

表1 Chi70 几丁质酶对Bt HD-73 增效作用Table 1 The synergistic activity of chitinase Chi70 to Bt HD-73
3 结论与讨论
Morgan(2001)等发现,在嗜线虫致病杆菌X.nematophilus PMFI296 毒素基因xptA1 上游有一个编码几丁质酶的基因序列,该基因同样编码648个氨基酸;推断该几丁质酶基因与其他毒素基因形成一个共同的致病基因簇,相互协同,共同对昆虫产生致病作用,但具体分子致病机制还需进一步研究。Chen(1996)等针对5种嗜线虫致病杆菌属和发光杆菌属线虫共生菌几丁质酶进行研究认为几丁质酶普遍存在于昆虫病原线虫共生菌中,但活性存在明显差异,几丁质酶活性的差异与其来源的菌株及测定方法有一定关联。
为了深入研究昆虫病原线虫共生菌中几丁质酶的特性,本研究对嗜线虫致病杆菌HB310 菌株中的几丁质酶基因chi70 进行了克隆、表达和分子生物信息学分析,分析结果表明chi70 基因全长1947 bp,编码648个氨基酸。生物信息学分析还发现Chi70 蛋白不含信号肽,但由于蛋白分泌机制具有多样性,这并不能证明它就是一种胞内酶,还需要其他方法进行进一步研究证实。
昆虫体壁、围食膜对环境及外界生物的侵害起物理和化学屏障作用,而其中起主要作用的组分是几丁质,几丁质成为害虫防治的突破点之一(Wiwat et al.,2000)。大量研究表明,几丁质酶与Bt 制剂混合使用可以提高 Bt 杀虫效果(Smirnoff,1971;Regev et al.,1996;Wiwat et al.,2000)。几丁质酶的来源非常广泛,Gopakakrishnan等(1995)把烟草天蛾几丁质酶基因转入到苜蓿银纹夜蛾核型多角体病毒AcMNPV 中,并成功表达了该酶;王芳表达了杆状病毒HaSNPV 几丁质酶基因并研究了表达蛋白对Bt 的增效作用(2004);丁学知等将烟草几丁质酶基因和苏云金芽胞杆菌cry1Ac 进行了双价基因的表达,这些研究都表明来源于不同物种的几丁质酶都可以增强Bt 的杀虫效果。本研究结果也显示来自嗜线虫致病杆菌HB310 菌株的几丁质酶基因重组表达的Chi70 蛋白对Bt HD-73 具有明显的增效作用。
嗜线虫致病杆菌中的几丁质酶在致病过程中起什么作用尚不清楚,Morgan 等认为,几丁质酶基因与其他致病基因形成一个基因簇,多种表达产物共同作用造成昆虫患病死亡。本研究对共生菌几丁质酶基因进行分析,初步了解其理化性质;通过原核表达体系对几丁质酶进行研究;并通过生测实验确定其增效作,为进一步研究几丁质酶Chi70 奠定基础,为下一步深入了解共生菌的作用机制提供理论依据。
References)
Arora N,Ahmad T,Rajagopal R,et al.A constitutively expressed 36 ku exochitinase from Bacillus thuringiensis HD-1[J].Biochem.Biophysi.Res.Communi.,2003,307:620-625.
Ausubel FM,Brent R,Kingston RE,et al.Short Protocols in Molecular Biology[M].Wiley,2002.
Chen G,Zhang Y,Li J,et al.Chitinase activity of Xneorhabdus and Photorhabdus species,bacterial associates of Entomopathogenic nematodes[J].Journal of Invertebrate Pathology,1996,68,101-108(1996).
Chaston JM,Suen G,Sarah LT,et al.The entomopathogenic bacterial endosymbionts Xenorhabdus and photorhabdus convergent lifestyles from divergent genomes[J].PloS ONE,2011,6(1):e27909.
Frank H,Jessica E,Schmitz,et al.Enrichment of chitinolytic microorganisms:isolation and characterization of a chitinase exhibiting antifungal activity against phytopathogenic fungi from a novel Streptomyces strain[J].Appl Microbiol Biotechnol,2005,66(4):4 34-42.
Gopalakrishnan B,Muthukrishnan S,Kramer KG.Baculovirus-mediated expression of a Manduca sexta chitinase gene:Properties of the recombinant protein[J].Insect Biochem.Mol.Biol.,1995,25(2):255-265.
Guo YJ,Experimental Technique of Protein Electrophoresis[M].Beijing:Science Press.2001,54-157[郭尧君,蛋白质电泳实验技术.北京:科学出版社.2001,54-157]
Li H,Liu KQ,Wang G.Measuring the chitinase activity of Trichoderma spp.by oxidation-reduction process[J].Journal of Zhongkai Agrotechnical College,2003,16(1):19-22.[李华,刘开启,王革.利用还原糖法测定木霉菌产几丁质酶特性[J].仲恺农业技术学院学报[J].2003,16(1):19-22]
Li XH,Wang QY,Lu XJ,et al.Susceptivty of different host insects to symbiotic bacteria of entomopathogenic nematodes[J].Entomological Knowledge,2004,40(1):39-42[李秀花,王勤英,陆秀君,等.不同昆虫寄主对昆虫病原线虫共生菌的敏感性比较[J].昆虫知识,2004,40(1):39-42]
Liu YZ,Luo CP,Liu YF,et al.Cloning and expression of a chitinase gene from Serratia marcescens strain C8-8[J].Agricultural Biotechnology,2014,2(3):56-59.
Morgan JAW,Sergeant M,Ellis D,et al.Sequence analysis of insecticidal genes from Xenorhabdus nematophilus PMFI296[J].Applied and Environmental Microbiology,2001,67(5):2062-2069.
Naosekpam S,Rajni V,Shanmugam V.Extracellular chitinases of fluorescent pseudomonads antifungal to Fusarium oxysporum f.sp.Dianthi causing carnation wilt[J].Curr.Microbiol.,2006,52:310-316.
Regev A,Keller M,Strizhov N,et al.Synergistic activity of a Bacillus thuringiensis δ-endotoxin and a bacterial endochitinase against Spodoptera littoralis larvae[J].Appl.Envir.Microbiol,1996,62(10):3581-3586.
Sanbrook J,Russell DW.(Translated by Huang PT).Molecular Cloning(3rdEdition)[M].Beijing:Science Press,2003.[Sanbrook J,Russell DW.(黄培堂译).分子克隆(第三版)[M].北京:科学出版社,2003]
Smirnoff WA.Effect of chitinase on the action of Bacillus thuringiensis[J].Can.Entomologist,1971,103:1829-1831.
Wang F,Gu QW,Wu JC,et al.Expression of HaSNPV chitinase gene in E.coli and insect cells and synergism of expressed products with Bt in killing insects[J].Chinese Journal of Biochemistry and Molecular Biology,2004 20(6):792~797
Wang QY,Nangong ZY,Lu XJ,et al.Purification of insecticidal proteins from Xenorhabdus nematophila HB310 and detection of their insecticidal activity[J].Acta Entomologica Sinica,2005,48(3):353-358[王勤英,南宫自艳,陆秀君,等.嗜线虫致病杆菌HB310 菌株杀虫蛋白的纯化及活性鉴定[J].昆虫学报,2005,48(3):353-358]
Wang QY,Nangong ZY,Yang J,et al.Toxic activity of a protein complex purified from Xenorhabdus nematophila HB310 to Plutella xylostellalarvae[J].Insect Science,2012,19:329-336.
Wiwat C,Thaithanun S,Pantuwatana S,et al.Toxicity of chitinase-producing Bacillus thuringiensis ssp.HD-1 toward Plutella xylostella[J].Invertebr.Pathol.,2000,76(4):270-277.
Zhang W,Ni L,Chen LY,et al,Optimizing conditions of chitinase production from Paecilomyces lilacinus strain FZ20289[J].Journal of Fujian Agriculture and Forestry University(Natural Science Edition),2006,34(2):195-199.[张雯,倪莉,程丽云,等.淡紫拟青霉FZ2 0289 产几丁质酶的条件优化[J].福建农林大学学报(自然科学版),2006,34(2):195-199]
Zhang ZX,Xu HH,Cheng MD.Calculating toxicity regression with EXCEL[J].Entomological Knowledge,2002,39(1):67-70[张志祥,徐汉虹,程东美.EXCEL 在毒力回归计算中的应用[J].昆虫知识,2002,39(1):67-70]
猜你喜欢
今日农业(2022年14期)2022-09-15 01:44:26
生物技术通报(2021年4期)2021-05-14 06:01:28
小学生作文(低年级适用)(2020年10期)2020-11-10 09:12:12
中国建筑装饰装修(2020年6期)2020-07-10 09:41:16
福建基础教育研究(2019年2期)2019-09-10 07:22:44
福建基础教育研究(2019年2期)2019-05-28 08:39:49
中成药(2018年2期)2018-05-09 07:20:04
中国酿造(2017年8期)2017-09-03 06:20:01
浙江大学学报(农业与生命科学版)(2017年1期)2017-04-17 07:05:02
天然产物研究与开发(2016年6期)2016-06-05 10:29:30
