
B 型烟粉虱在甘蓝上的产卵行为观察
2014-12-16 01:57:30张晓曼
环境昆虫学报 2014年6期
张晓曼,王 甦,罗 晨,张 帆
(北京市农林科学院植物保护环境保护研究所,北京 100097)
烟粉虱Bemisia tabaci(Gennadius)隶属同翅目粉虱科小粉虱属,是世界范围内重要的农作物入侵性害虫(De Barro et al.,2011),它可通过刺吸植物汁液,分泌蜜露和传播病毒等多种方式危害植物,是大田作物,蔬菜和园林植物上的重要害 虫(Brown and Bird,1992;Perring et al.,1993;EPPO,2004)。烟粉虱与温室白粉虱Trialeurodes vaporariorum 常常混合发生,且体型微小,形态特征相似,很难区分(褚栋等,2005)以往报道烟粉虱与温室白粉虱产卵方式有所不同,温室白粉虱在光滑的叶片上多将卵排列成圆形或半圆形,在多毛叶片上多散产,而烟粉虱通常为散产(吴杏霞,胡敦孝,2000;Hoddle,2011)。烟粉虱具有多种种下生物型(隐种)其中B 型(MEA1)烟粉虱也称银叶粉虱B.tabaci(=Bemisia argentifolii)为世界性危害害虫(Bellows et al.,1994)。仅以巴西的圣保罗为例,目前危害蔬菜的烟粉虱只有B 型烟粉虱(Kelly et al.,2011)。为了更好的防治烟粉虱,人们开展了大量的基础生物学及生理学研究。其中对于烟粉虱繁殖行为及繁殖策略的研究,可以帮助我们认识烟粉虱的及传行为播规律,提高对烟粉虱的专性防治效率(Liu et al.,2007)。
在研究烟粉虱生物学习性及与温室白粉虱的区别时,曾有研究报道烟粉虱产卵方式为多散产,罕见为弧形、圆或半圆(吴杏霞,胡敦孝,2000;朱国仁等,2006;Hoddle,2011),Hoddle 介绍(http:∥biocontrol.ucr.edu/bemisia.html)(2011),而温室白粉虱通常产卵的排列方式为圆形或半圆。昆虫的产卵策略与营养供应和天敌侵扰有很大相关性(Rouault et al,2007),已有研究表明烟粉虱的捕食性天敌小黑瓢虫Delphastus catalinae 最喜欢在甘蓝上取食烟粉虱(罗宏伟,2010)。而烟粉虱在相同的雌虫密度下在甘蓝上的产卵量与其它植物相比产卵量最多(林克剑,2008),所以表明烟粉虱为了平衡天敌对其在甘蓝上较多的捕食量而会选择增加产卵量。烟粉虱的产卵策略在不同寄主植物间有较明显的差异。本文表明在甘蓝上少散产,多将卵排列成弧形、圆或半圆形,当烟粉虱将卵以 弧形、圆或半圆形的产卵方式排列时,边取食边产卵,不论从能量来源,体力消耗还是节约空间的角度来考虑,都有利于烟粉虱增加其产卵量,来平衡天敌对其的捕食。
1 材料与方法
1.1 虫源和供试植物
粉虱采自北京市农林科学院温室内由甘蓝Brassica oleracea L.var.caqitata 繁殖,寄主植物种植在温室内(温度25℃-30℃,湿度RH 50 %-60 %,光照L∶D=16 h∶8 h),品种为秋甘一号F1(北京市农林科学院蔬菜研究所提供),栽培土为富养基质,种植时,将基质加适量清水,搅拌均匀,分装在在上口径20 cm 的塑料花盆内,当苗高15 cm 时供试。
1.2 粉虱的鉴定
1.2.1 用体式显微镜和倒置显微镜观察粉虱的形态和复眼
取正在产卵(卵排列方式为圆形)的粉虱浸泡在2 mol/L NaOH 碱性溶液中,55℃水浴2 h,在Nikon ECLiPSE Ti 倒置显微镜下观察粉虱复眼的形态结构(目镜10×、物镜40×)(褚栋等,2008)。
1.2.2 DNA 条形码技术鉴定烟粉虱的生物型
烟粉虱mt COⅠ由通用引物扩增并测序,三个B 型和两个Q 型烟粉虱的mt COⅠ基因序列从网站 NCBI 中收集(http:∥www.ncbi.nlm.nih.gov/),与我们实验室内粉虱得到的8个基因序列在网站上用Multalin(http:∥multalin.toulouse.inra.fr/multalin/multalin.html)进行比对,系统进化树的构建利用MEGA 4 和Clustalx 软件完成(Kumar et al.,2004),采用邻接法基于“bootstrap”函数构建邻接(Neighbor-joining)系统树。
1.3 B 烟粉虱产卵排列方式
寄主植物甘蓝在长宽高各为35 cm 的120 目纱网养虫笼内养殖,当甘蓝苗高15 cm,选择叶片面积大约为100 cm2的叶片,用纱网罩住,从10 株棉花上选择10个合适的叶片,在纱网内释放B 型烟粉虱,平均8 对/cm2,每个纱网罩内均放入800 对烟粉虱,此时定义为F0代,同样的方法再连续两代释放烟粉虱分别定义为F1代和F2代每代选择叶片重复次数为10。分别接烟粉虱5 d 后记录每个叶片上B 型烟粉虱不同产卵排列形状(圆形,半圆和弧形)的数量,两个卵之间的距离不超过0.01 mm 且卵的排列方式趋于圆形时,记录为圆形排列的卵,当第一个卵与最后一个卵之间处在圆的直径上,且处在半圆上卵与卵之间的距离不超过0.01 mm 记录为半圆,当第一个卵与最后一个卵之间的距离小于圆的直径距离时,记录为弧形(0<弧形<π)。
1.4 数据处理与分析
利用SAS16.0 对数据间进行比较分析,采用ANOVA 进行方差分析,采用LSD 法进行平均数之间的比较,P<0.05。
2 结果与讨论
通过显微镜与倒置显微镜的观察,前翅呈较明显的屋脊状,且上下部复眼有链接,判断为烟粉虱(图1A),且通过线粒体CO I 基因的鉴定和进化树的构建确定实验室内的烟粉虱为B 型烟粉虱。B 型烟粉虱在甘蓝上的产卵排列方式大部分为圆形,弧形和半圆形,少散产,烟粉虱产卵时以取食点为中心,身体为半径,转动身体,边取食边产卵,从而卵的排列方式为圆形,弧形和半圆(图1:BC)。
连续观察三代烟粉虱的产卵排列方式,从表1、表2、表3 中可以看出,三代B 烟粉虱产卵的排列形状以圆形和弧形为主,少散产,数据显示三代B 型烟粉虱在甘蓝叶片上产卵的排列形状是以圆形,半圆形和弧形为主,少散产,所占比例分别为圆形45.1 %、弧形46.9 %、半圆8.0 %,圆形和弧形占92.0 %。第一代烟粉虱的产卵排列方式比例为圆形:弧形:半圆形=1∶1.06∶0.11,第二代烟粉虱产卵排列方式比例为圆形∶弧形∶半圆形=1∶0.86∶0.17,第三代烟粉虱产卵排列方式比例为圆形∶弧形∶半圆形=1∶1.20∶0.25。通过spss16.0 中LSD 法检验三代烟粉虱产卵排列方式比较,在每一代中,圆形和弧形没有显著性差异,但都与半圆有显著性的差异,F0、F1和F2代的相同的烟粉虱卵排列方式如:圆形、弧形和半圆形三代中均没有显著性差异。

表1 F0代B 型烟粉虱在甘蓝叶片上产卵排列方式的对比Table 1 Comparison of oviposition arrangement of F0generation Bemisi argentifolii(type B B.tabaci)on cabbage leaves
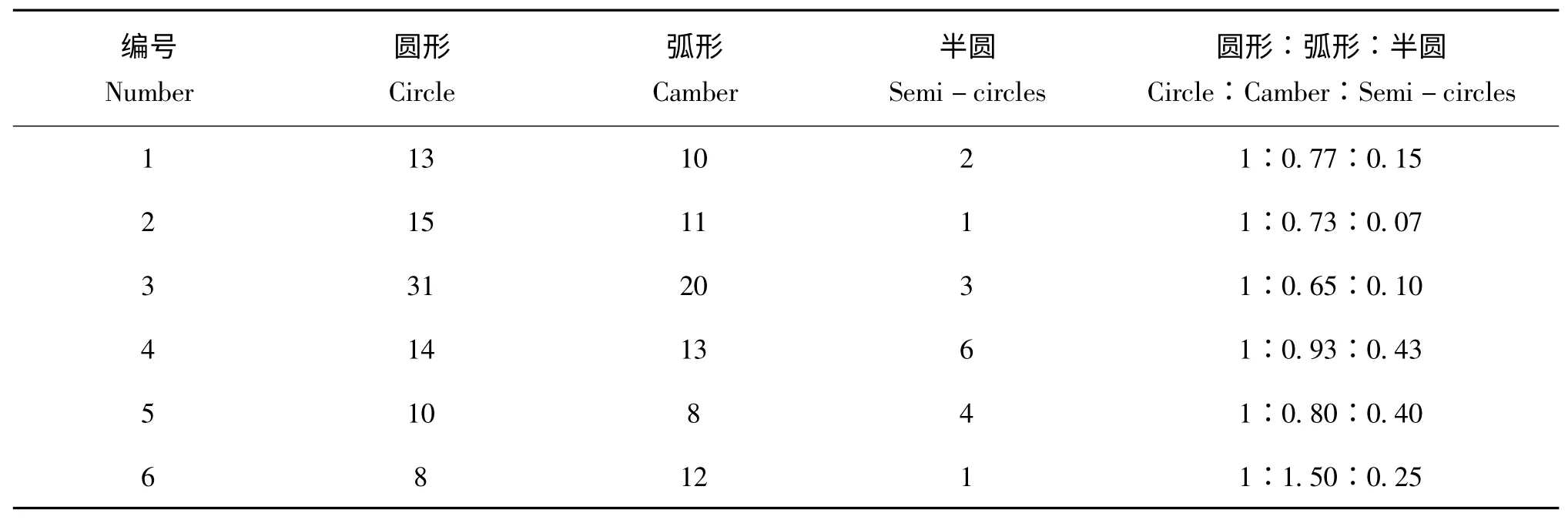
表2 F1代B 型烟粉虱在甘蓝叶片上产卵排列方式的对比Table 2 Comparison of oviposition arrangement of F1generation Bemisia argentifolii on the cabbage leaf
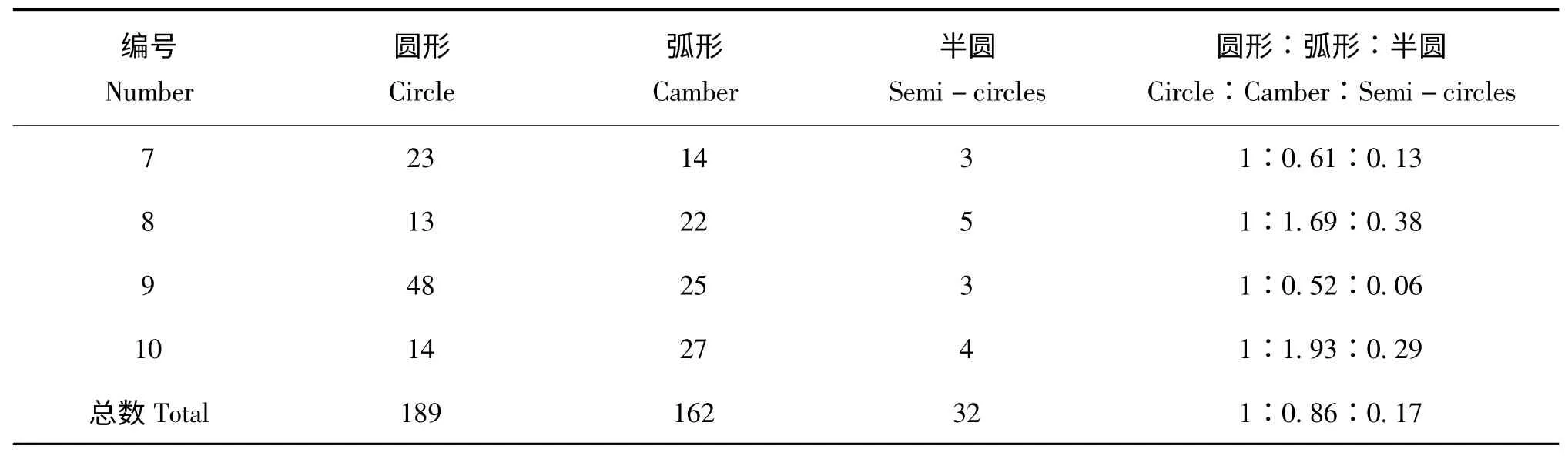
(续上表)

表3 F2代B 型烟粉虱在甘蓝叶片上产卵排列方式的对比Table 3 Comparison of oviposition arrangement of F2generation Bemisia argentifolii on the cabbage leaves
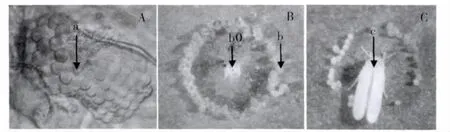
图1 A,烟粉虱复眼,箭头a 为上下部复眼连接处;B,箭头b0 为烟粉虱取食原点;C,箭头c 为正在产卵的烟粉虱Fig.1 A,Compound eye of B.tabaci,arrows“a”is the junction of up and down compound;B,arrows“b0”is feeding origin;C,arrows“c”is that B.tabaci was laying eggs.

表4 三代烟粉虱产卵排列方式的比较(mean±SE)(F2,27)Fig 4 Comparison of oviposition arrangement of three generation Bemisia tabaci
3 结论与讨论
昆虫的产卵行为和产卵策略与营养供养和天敌的侵扰压力有很大的关系(Rouault et al,2007)。B 型烟粉虱在保证子代食物源的情况下,可能以此种产卵排列方式增加单位面积的产卵量来抗衡天敌对其种群的抑制。
通过我们的观察,B 型烟粉虱在多毛的棉花叶片上多散产,偶尔卵也会排列成圆形或半圆,在光滑的甘蓝叶片上产卵时把口器插入叶片,以吸食点为圆心,转动身体产卵,卵多数排列成圆形,弧形或半圆,所占比例分别为圆形45.1 %、弧形46.9 %、半圆8.0 %,圆形和弧形占92.0 %。其在小白菜和快菜上产卵方式也是如此,区别于前人的研究认为烟粉虱将卵散产在叶片背面(吴杏霞,2000,朱国仁,2006),也不同于国外研究人员Hoddle(2011)对B 型烟粉虱的介绍(http:∥biocontrol.ucr.edu/bemisia.html),其描述B 型烟粉虱将卵散乱的产在叶片背面,而温室白粉虱产卵时将卵排列成圆形或半圆形。本次试验结论与前人研究不同的原因可能是由于环境的影响。
有研究表明,B 型烟粉虱在甘蓝、西葫芦和辣椒上的刺吸取食波形反应,其在甘蓝上的适合度是最高的,60 min 内留存B 型烟粉虱的百分比为100 %(岳梅等,2006),林克剑的研究表明,B 型烟粉虱在5.0 头/cm2雌虫密度条件下产卵量比较为甘蓝>苘麻>黄瓜>烟草>棉花(林克剑,2008),对比之下其在甘蓝上的产卵量最高。本实验结果表明B 型烟粉虱产卵时以取食点为中心,挪动身体将卵排列成圆形,半圆或弧形,边取食边产卵,这个结果与岳梅研究烟粉虱在甘蓝上的适合度最高一致,烟粉虱在甘蓝上采用这种产卵策略,在相同的时间内,与其它散产叶片上的烟粉虱相比增加了取食量,且卵是紧密排列的,节约了空间,所以这可能是B 型烟粉虱在相同密度下产卵量最高的一个主要原因。
B 型烟粉虱的这种产卵排列方式也有可能是防御捕食性天敌的一种策略,B 型烟粉虱的这种产卵排列方式增加了产卵量,在同样满足捕食性天敌取食的情况下,这种产卵排列方式与散产相比能够防止更多的卵不被天敌取食。也有研究表明,烟粉虱的捕食性天敌小黑瓢虫Delphastus catalinae最喜欢在甘蓝上取食烟粉虱,而不喜欢在番茄和茄子上取食烟粉虱(罗宏伟,2010)。所以烟粉虱在甘蓝上以取食点为中心,身体为半径转动取食,节约时间节约体力节约空间增加产卵量来平衡捕食性天敌对其的大量捕食。也有一种可能跟叶片有无叶毛有关,在有毛的叶片上烟粉虱卵散产,相对产卵量低于甘蓝叶片上的产卵量,同时捕食性天敌在无毛叶片上的捕食量也增加,这样便达到了一种供需平衡,如小黑瓢虫在无毛的一品红Annetle Hegg Brilliant Diamond 品种相对于多毛的Lilo 品种上捕食量和产卵量都有增加(Heinz,1994)。
通过我们的观察发现,B 型雌性烟粉虱在甘蓝叶片上产卵时,仍有少部分雄性的烟粉虱继续停留在雌性烟粉虱旁,随着雌性B 型烟粉虱产卵一起转动(卵不散产时)将卵排列成圆形,半圆或弧形。B 型烟粉虱产卵时以吸食点为圆心产卵时,遇到叶脉、其它烟粉虱产的卵或其它障碍时,烟粉虱便会离开其产卵的地方,卵便会排列成半圆或弧形,或者是其自身的生理因素,而改变产卵的位置,所以烟粉虱产卵排列方式是烟粉虱自身生理因素与外界环境的共同作用结果。通过对烟粉虱产卵行为的研究,对其种群发展及其防止天敌对其种群的抑制的策略有潜在的意义。
References)
Bellows TSJr,Perring TM,Gill RJ,et al.Description of a species of Bemisia(Homoptera:Aleyrodidae)[J].Ann.Entomol.Soc.Am,1994,87,195-206.
Brown JK,Bird J.Whitefly-transmitted geminiviruses and associated disorders in the Americas and the Caribbean Basin[J].Plant Disease,1992,76,220-225.
Chu D,Wang B,Zhang SH,et al.Rapid differentiation between Bemisia tabaci and Trialeurodes vaporariorum adults by microscopic examination of compoundeyes[J].Chinese Bulletin of Entomology,2008,45.(1):154-155.[褚栋,王斌,张四海等.一种快速鉴别烟粉虱与温室白粉虱成虫的方法-复眼镜检法[J].昆虫知识,2008,45(1):154-155]
Chu D,Zhang YJ,Cong B,et al.Sequence analysis of mtDNA COI gene and molecular phylogeny of different geographical populations of Bemisia tabaci(Gennadius)[J].Scientia Agricultura Sinica,2005,38(1):76-85.[褚栋,张友军,丛斌等.烟粉虱不同地理种群的mtDNACOI 基因序列分析及其系统发育[J].中国农业科学,2005,38(1):76-85]
De Barro Paul J,Liu SS,Boykin LM,et al.Bemisia tabaci:A statement of species status[J].Annu.Rev.Entomol.,2011,56:1-19.
EPPO,Bemisia tabaci.Bulletin OEPP/EPPO[J].Bulletin,2004,34:281-288.
HebertPDN,Ratnasingham S,de Waard JR.Barcoding animal life:cytochrome c oxidase subunit I divergences among closely related species[J].Proc.R.Soc.Biol.Sci.Ser.B,2003,270:S96-S99.
Heinz KM,Parrella MP.Poinsettia(Euphorbia pulcherrima Willd.Ex koltz.)cultvar medizted differences in performance of five natural enemies of Bemisia argentifolii bellows and perring,n.sp(Homoptera:Aleyrodidae)[J].Biol.Control,1994,4(4):305-318.
Mark H.Thebiology and management of the Silverleaf Whitefly,Bemisia argentifolii bellows and Perring(Homoptera:Aleyrodidae)on greenhouse grown ornamentals[J].Extension Specialist and Director of Center for Invasive Specie Research,2011,
Kelly CGR,Julio MM,Jesús NC,et al.Only the B biotype of Bemisia tabaci is present on vegetables in São Paulo State[J].Brazil.Sci.Agric.(Piracicaba,Braz.),2011,6(1):120-123.
Kumar S,Tamura K,Nei M.MEGA3:integrated softwarefor molecular evolutionary genetics analysis and sequencealignment[J].Briefings in Bioinformatics,2004,5:150-163.
Lin KJ,Wu KM,Zhang YJ,et al.The feeding and oviposition behaviors of Bemisia tabaci(Gennadius)biotype B on five host plants[J].Acta Phytophylacica Sinica,2008,35(3):199-204.[林克剑,吴孔明,张永军,等.B 型烟粉虱成虫对五种寄主植物的取食和产卵行为[J].植物保护学报,35(3):199-204]
Liu SS,De Barro PJ,Xu J,et al.Asymmetric mating interactions drive widespread invasion and displacement in a whitefly[J].Science,2007,318:1769-1772.
Luo C,Xiang YY,Guo XJ,et al.Comparative on development and reproduction between Bemisia tabaci biotype B and Trialeurodes vaporariorum on four species of host-plants[J].Acta Ecologica Snice,2007,27(3):1035-1040.[罗晨,向玉勇,郭晓军,等.寄主植物对B 型烟粉虱(Bemisia tabaci)和温室粉虱(Trialeurodes vaporariorum)个体发育和种群繁殖的影响[J].生态学报,2007,27(3):1035-1040]
Luo HW,Wang ZH,Wang LD,et al.Effects of host plants on development,survival and reproduction of Delphastus catalinae(Horn),apredatorof Bemisia tabaci(Gennadius)[J].Journal of Fujian Agriculture and Forestry University(Natural Science Edition),2010,39(3):231-235.[罗宏伟,王竹红,王联德,等.寄主植物对烟粉虱捕食性天敌-小黑瓢虫发育、存活和繁殖力的影响[J].福建农林大学学报(自然科学版),2010,39(3):231-235]
Perring TM,Cooper AD,Rodriguez,et al.Identification of a whitefly species by genomic and behavioural studies[J].Science,1993,259,74-77.
Ren SX,Qiu BL,Ge F,et al.Research progress of the monitoring,forecast and sustainable management of whitefly pests in China[J].Chinese Journal of Applied Entomology,2011,48(1):7-15.[任顺祥,邱宝利,戈峰,等.粉虱类害虫的监测预警与可持续治理技术透视[J].应用昆虫学报,2011,48(1):7-15]
Rouault G,Battisti A,Roques A.Oviposition sites of the cypress seed bug Orsillus maculates and response of the egg parasitoid Telenomus gr.floridanus[J].BioControl,2007,52:9-24.
Wu XX,Hu DX.The identification of Bemisia tabaci,Trialeurodes vaporariorum and silverleaf whitefly[J].Beijing Agricultural Sciences,2000,36-40.[吴杏霞,胡敦孝.温室白粉虱、烟粉虱及银叶粉虱的识别.北京农业科学[J].北京农林业科学,2000,36-40]
Wu XX,Hu DX.The difference between Bemisia tabaci and Trialeurodes vaporariorum[J].Plant Protection,2001,27(5):15-18.[胡敦孝,吴杏霞.烟粉虱和温室白粉虱的区别[J].植物保护,2001,27(5):15-18]
Yue M,Luo C,Guo XJ,et al.Probing and feeding behaviors of the whitefly Bemisia tabaci(Gennadius)B.biotype on three host plants[J].Acta Entomologica Sinica,2006,49(4):625-629.[岳梅,罗晨,郭晓军,等.B 型烟粉虱在甘蓝、西葫芦和辣椒上的刺吸取食行为[J].昆虫学报,2006,49(4):625-629]
Zhu GR,Wu QJ,Zhang YJ,et al.Identification and control of two kinds of whitefly on vegetables[J].China Vegetables,2006,(6):49-51.[朱国仁,吴青君,张友军等.蔬菜上两种粉虱的识别与防治[J].中国蔬菜,2006,6:49-51]
猜你喜欢
小学生学习指导(高年级)(2023年10期)2023-10-28 09:00:46
昆虫学报(2022年11期)2022-12-28 02:44:18
孩子(2021年1期)2021-01-27 02:24:03
上海农业科技(2020年4期)2020-08-19 11:33:26
植物保护(2018年5期)2018-12-05 10:36:16
小学生学习指导(高年级)(2018年10期)2018-10-10 01:01:50
中等数学(2018年6期)2018-08-02 02:07:22
新农业(2017年2期)2017-11-06 01:02:23
新疆农垦科技(2016年10期)2016-06-15 20:29:33
生物灾害科学(2015年3期)2015-03-11 16:38:37
