
表达猴免疫缺陷病毒Env蛋白的重组鸡痘病毒的构建和筛选
2014-10-27 09:04朱羿龙李昌刘存霞杜寿文王茂鹏叶飞谭鹏邢彬刘继朱光泽郭焱金宁一
生物技术通讯 2014年3期
朱羿龙,李昌,刘存霞,杜寿文 ,王茂鹏,叶飞,谭鹏,邢彬,刘继,朱光泽,郭焱,金宁一
1.军事医学科学院 军事兽医研究所,吉林 长春 130122;2.长春中医药大学,吉林 长春 130117
据世界卫生组织统计,2010年新感染人免疫缺陷病毒(HIV)的患者约270万人,而截至2010年底,约有3400万人携带HIV,所以迫切需要安全、有效的疫苗来应对HIV的蔓延。但是,由于抗原表位的时常突变、免疫原性弱,以及疫苗所诱导的抗原免疫反应时间短等原因,导致疫苗研发屡遭挫败[1-3]。对于其他病毒传染病,如麻疹、腮腺炎和风疹等,这种类似于免疫原性的问题是通过发展减毒疫苗株来解决的[4],但是对于HIV和猴免疫缺陷病毒(SIV)而言,减毒活疫苗虽然很具有吸引力[5-6],但却存在前病毒整合和某些情况下转变为野生毒株的风险[7]。相反,活病毒在作为HIV疫苗载体时具备了相当优秀的免疫原性[8-11],其中以禽痘病毒为载体的HIV疫苗已开始临床应用,如Aventis Pasteur公司开发的重组金丝雀病毒载体ALVAC与gp120亚单位疫苗联合应用,并均可在受试者中检测到CTL反应[12]。
SIV与HIV-1具有一定的同源性,部分SIV毒株接种于亚洲猴子,会使其感染并产生类似艾滋病的症状[13],因此,用SIV感染亚洲猴属作为模型被推荐替代HIV-1感染,使得SIV作为HIV的模式病毒在疫苗领域得到广泛应用。包膜糖蛋白(Env)在病毒进入宿主细胞的过程中起着至关重要的作用,我们以SIV env作为目的基因,以中国鸡痘病毒(fowlpox virus,FPV)疫苗株FPV282E4为载体,以EGFP为标记基因,采用挑斑方法筛选重组FPV,利用PCR、RT-PCR和Western印迹进行鉴定和遗传稳定性分析,最终获得一株稳定表达SIV Env蛋白的重组FPV,通过其所产生的免疫应答反应,可为HIV疫苗研究提供可资借鉴的数据。
1 材料和方法
1.1 材料
含有SIV env基因的质粒pVR-SIV env由中国疾病预防控制中心病毒病预防控制所冯霞博士惠赠;穿梭质粒 pTKET[14]、大肠杆菌 DH5α、FPV282E4株由本实验室保存;8~10日龄SPF鸡胚购自北京梅里亚维通实验动物技术有限公司,用于制备鸡胚成纤维细胞(CEF)。
1.2 重组质粒的构建
以pVR-SIV env为模板设计扩增SIV env的引物 F(5'-GCCACCATGGGATGCCTGGGAAAC-3',添加KpnⅠ酶切位点及kozak序列)和R(5'-TCACTG CCTCAGCTTAGCCAG-3',添加SalⅠ酶切位点)。反应条件:94℃变性 30 s;60℃退火 30 s,72℃延伸130 s,30个循环,72℃延伸10 min。将PCR产物连接到pMD18-T载体上,酶切鉴定正确后进行DNA测序。用KpnⅠ与SalⅠ双酶切T载体上测序正确的目的基因片段,插入FPV穿梭载体pTKET的相应位点,构建重组FPV质粒pTKET-SIV env(图1)。
1.3 FPV的重组、筛选和纯化
以5MOI(感染复数)的FPV282E4感染生长至80%融合的CEF,37℃、5%CO2条件下培养2~3 h,用QIAGEN公司的转染试剂Effectene Transfection Reagent将重组质粒转染至CEF中,继续培养72 h,在荧光显微镜下观察绿色荧光后,收取病变细胞并超声波破碎,将其接种于单层CEF中培养72 h,在荧光显微镜下挑取蚀斑,经超声波破碎,再次接种于单层CEF中,重复上述操作,待形成的细胞病变处无非绿色荧光病变细胞时,挑取噬斑进行扩增、鉴定。
1.4 重组FPV的整合与转录
将纯化的重组FPV以5MOI接种CEF,在37℃、5%CO2条件下培养72 h后收集细胞,提取感染细胞的总基因组和RNA,将RNA反转录成cDNA,以基因组和cDNA为模板,扩增SIV env、FPV-P4b(上游引物为5'-GGACGCGTATTGATTCACACCGTATTA CAGAGG-3',下游引物为5'-CGCCCGGGTTCTCC TAATAAGTTACACCGTTTG-3')、FPV-TK(上游引物为5'-GGACGCGTCAGCAGGTGCTAAACAACAA-3',下游引物为5'-GGCTGCAGCGGTAGCTTAACG CCGAATA-3')基因。反应条件:94℃变性30 s;60℃退火30 s,72℃延伸130 s,30个循环;72℃延伸10 min。TK基因作为VAVC的一个常用插入位点,也用于FPV和其他禽痘病毒的重组位点[15-16],因此选取TK基因作为重组FPV是否筛纯的鉴定指标。P4b基因存在于所有FPV中,通常将其用于FPV的特异性鉴定[17]。
1.5 重组FPV表达产物的检测
将纯化的重组FPV以5MOI接种CEF,在37℃、5%CO2条件下培养72 h后收集细胞,用裂解缓冲液RIPA裂解细胞,离心取上清,加入5×SDS-PAGE缓冲液,沸水浴10 min,经SDS-PAGE后转移到Im⁃mun-Blot PVDF 膜上,以兔抗 gp120/160(Mac293)(购自Eneyme公司)和鼠抗GAPDH抗体(购自碧云天公司)为一抗,辣根过氧化酶标记的山羊抗兔IgG(购自中杉金桥公司)和辣根过氧化酶标记的山羊抗鼠IgG(购自中杉金桥公司)为二抗进行抗体结合反应,利用ECL方法进行蛋白表达分析,设立GAPDH为内参。
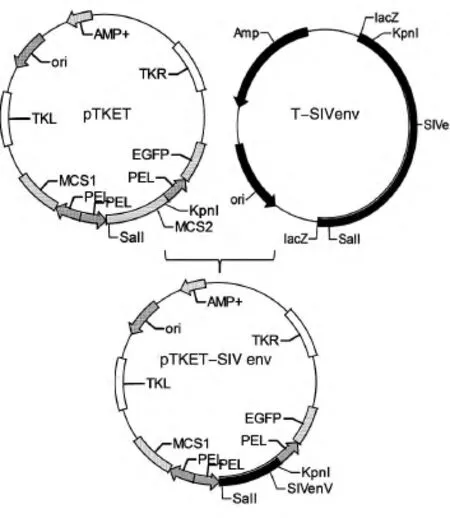
图1 重组鸡痘病毒穿梭质粒的构建
1.6 遗传稳定性分析
将筛选纯化的重组FPV在CEF中连续传代20次,分别选取传代次数为1、5、10、15、20的感染病毒的细胞,观察荧光并提取其基因组、总RNA和总蛋白,利用PCR、RT-PCR、Western印迹检测SIV env基因在重组FPV中的遗传稳定性。
1.7 重组FPV在哺乳动物细胞中的表达
将纯化的重组FPV以5 MOI接种BHK21细胞,在37℃、5%CO2条件下培养72 h后,在荧光显微镜下观察细胞能否表达荧光。
2 结果
2.1 重组质粒的构建及鉴定
按照前述方法扩增SIV env基因,可获得2.1 kb左右的目的片段,将其分别连接到pMD18-T载体中,经KpnⅠ和SalⅠ双酶切,可切出2.1 kb左右的目的片段,表明扩增成功。经测序证明为目的基因序列(图略)。
2.2 重组FPV穿梭质粒pTKET-SIV env酶切鉴定
重组FPV穿梭质粒pTKET-SIV env经KpnⅠ和SalⅠ酶切,可看到2.1 kb左右的目的片段及4.8 kb左右的载体片段,表明质粒构建成功(图2)。
2.3 重组FPV的整合与转录鉴定结果
提取重组FPV感染CEF的基因组和总RNA,进行PCR和RT-PCR,结果见图3,重组FPV基因组和cDNA均可扩增出SIV env片段(2160 bp)、P4b片段(578 bp),证明目的基因已整合到重组FPV中并能进行转录。由图3还可以看出,重组FPV基因组未扩增TK基因,而野生型毒株可以扩增出TK特异性条带,说明重组FPV已经获得纯化。
2.4 重组FPV表达产物的检测结果
将重组FPV感染CEF的裂解液行SDS-PAGE和转膜后,分别与兔抗gp120/160、鼠抗GAPDH抗体和辣根过氧化酶标记的山羊抗兔IgG、辣根过氧化酶标记的山羊抗鼠IgG反应,用ECL方法进行蛋白表达分析,以GAPDH为内参,结果见图4。重组病毒感染的细胞可检测出相对分子质量约120×103的SIV Env目的条带,而野生型FPV感染检测为阴性,用GAPDH抗体检测可见相对分子质量约40×103的目的条带,表明目的蛋白成功表达且具有抗原性。
2.5 遗传稳定性分析结果
将经挑斑纯化的重组FPV连续传代20次,分别取第1、5、10、15、20代病毒观察,可见每代均有荧光(图5);提取基因组和总RNA进行PCR和RT-PCR检测SIV env基因,结果表明各代重组FPV均能扩增出2160 bp的目的片段(图6);同时制备蛋白样品进行Western印迹,可见相对分子质量约120×103的特异性蛋白条带,而FPV检测为阴性(图7)。表明重组FPV传代20次内具有良好的遗传稳定性。
2.6 重组FPV在真核细胞中的表达情况
为了验证本研究构建的重组FPV能否在哺乳动物中表达外源蛋白,以5MOI病毒量感染BHK21细胞,72 h后观察,可以看到感染重组病毒的细胞有大量荧光表达,而感染野生毒株的对照细胞则无(图8),表明重组FPV能在哺乳动物细胞中表达。
3 讨论

图2 重组穿梭质粒的酶切鉴定
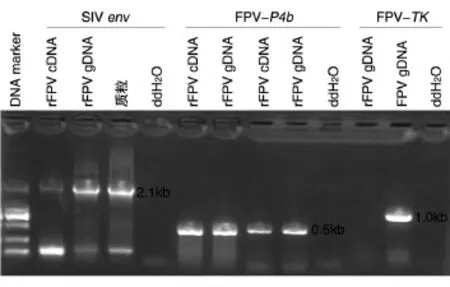
图3 重组FPV的PCR产物电泳图谱
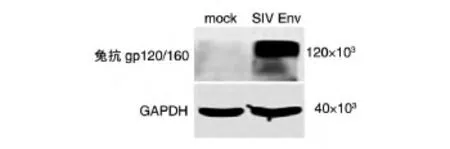
图4 重组FPV表达产物的Western印迹鉴定
艾滋病在1981年突然出现,并在极短的时间内迅速蔓延,至今已造成约6000万人感染,而且新感染者人数正以每年260万持续增长[18]。艾滋病已成为世界上最具破坏性的传染病之一,虽然各国都在尽力研究抗艾滋病药物,企图阻止病毒在不同感染阶段时的复制,但由于HIV基因在治疗期间的突变,使得药物往往无法达到理想的治疗效果。因此,研制安全和有效的疫苗,是控制、消除艾滋病流行的主要措施,近年来研究者已逐步开始关注以病毒为载体的HIV疫苗。
FPV载体是应用较为广泛的禽痘病毒。鉴于禽痘病毒具有严格的胞浆复制、忠实表达外源蛋白、自然条件下只感染禽类、流产性感染哺乳动物且不产生感染性子代病毒粒子等特点,且其具有很大的外源基因容量,构建相对容易,因此以FPV为载体的重组疫苗已经应用于哺乳动物和禽类的研究中[19-22]。
SIV与HIV具有很高的同源性。由于SIV的遗传结构和生物特性与HIV非常相似,同时SIV猕猴动物模型与HIV患者有许多相似之处[23],主要表现为身体消瘦、机会性感染及CD4+T淋巴细胞大量丢失,因此,SIV感染模型在目前最适于艾滋病发病机制及疫苗战略研究[24]。
图5 不同代次重组FPV的荧光分析
逆转录病毒的包膜糖蛋白在感染初期发挥着重要的作用,它涉及结合细胞膜和细胞表面受体使病毒融合进入细胞;在感染后期,Env在病毒装配的过程中也扮演重要角色,有证据表明包膜基质蛋白与Env在细胞内相互作用直接影响病毒粒子在极化上皮细胞的释放位置[25-26],位于胞内区的Env能够相互作用,同时还会影响Env的掺入和感染力。此外,去除细胞质域能够增加被感染细胞表面Env的表达量和掺入病毒样颗粒以及Env融合活性[27],同时Env还具备良好的免疫原性,能够有效刺激SIV特异性IFN-γ分泌细胞的产生和诱导较高水平的Env特异性反应[28],能够产生高浓度的特异性抗体[29],能够有效诱导产生CD4+T淋巴细胞增殖效应[30]。因此,Env作为有效抗原被广泛应用于疫苗研发。
本实验以SIV env为抗原基因,构建能稳定表达Env蛋白的重组FPV,并采用挑取噬斑的方法进行筛选,同时以EGFP基因为报告基因,使重组FPV在感染CEF后能特异性表达绿色荧光蛋白,由此大大提高重组病毒的检出率,又可避免用BrdU加压筛选导致基因突变的不利影响。用PCR、RT-PCT、Western印迹对重组FPV进行了鉴定和遗传稳定性分析,表明我们构建的重组FPV能同时表达SIV Gag和SIV Env,具备更广泛的免疫原性,且重组FPV能在BHK21细胞中大量表达,为其在哺乳动物中的应用及HIV疫苗研究提供了可资借鉴的数据。
图6 不同代次重组FPV的PCR产物电泳图谱
图7 不同代次重组FPV表达产物的Western印迹

图8 重组FPV在BHK21细胞中的表达
[1]Gallo R C.The end or the beginning of the drive to an HIV-preventive vaccine:a view from over20 years[J].Lancet,2005,366(9500):1894-1898.
[2]Saunders K O,Rudicell R S,Nabel G J.The design and evaluation of HIV-1 vaccines[J].Aids,2012,26(10):1293-1302.
[3]Burton D R,Ahmed R,Barouch D H,et al.A blueprint for HIV vaccine discovery[J].Cell Host Microbe,2012,12(4):396-407.
[4]Watson J C,Hadler S C,Dykewicz C A,et al.Measles,mumps,and rubella--vaccine use and strategies for elimina⁃tion of measles,rubella,and congenital rubella syndrome and control of mumps:recommendations of the Advisory Commit⁃tee on Immunization Practices(ACIP)[J].MMWR Recommend Rep,1998,47(RR-8):1-57.
[5]von Gegerfelt A S,Liska V,Li P L,et al.Rev-independent simian immunodeficiencyvirusstrainsare nonpathogenicin neonatal macaques[J].J Virol,2002,76(1):96-104.
[6]von Gegerfelt A S,Alicea C,Valentin A,et al.Long lasting control and lack of pathogenicity of the attenuated Rev-inde⁃pendent SIV in rhesus macaques[J].AIDS Res Human Retro⁃viruses,2006,22(6):516-528.
[7]Baba T W,Liska V,Khimani A H,et al.Live attenuated,multiply deleted simian immunodeficiency virus causes AIDS in infant and adult macaques[J].Nat Med,1999,5(2):194-203.
[8]Johnston R E,Johnson P R,Connell M J,et al.Vaccination of macaques with SIV immunogens delivered by Venezuelan equine encephalitis virus replicon particle vectors followed by a mucosal challenge with SIVsmE660[J].Vaccine,2005,23(42):4969-4979.
[9]Demberg T,Florese R H,Heath M J,et al.A replicationcompetent adenovirus-human immunodeficiency virus(Ad-HIV)tat and Ad-HIV env priming/Tat and envelope protein boost⁃ing regimen elicits enhanced protective efficacy against simian/human immunodeficiency virus SHIV89.6P challenge in rhe⁃sus macaques[J].J Virol,2007,81(7):3414-3427.
[10]Hansen S G,Ford J C,Lewis M S,et al.Profound early con⁃trol of highly pathogenic SIV by an effector memory T-cell vaccine[J].Nature,2011,473(7348):523-527.
[11]Robert-Guroff M.Replicating and non-replicating viral vec⁃tors for vaccine development[J].Curr Opin Biotechnol,2007,18(6):546-556.
[12]Pitisuttithum P,Rerks-Ngarm S,Bussaratid V,et al.Safety and reactogenicity of canarypox ALVAC-HIV(vCP1521)and HIV-1 gp120 AIDSVAX B/E vaccination in an efficacy trial in Thailand[J].PloS One,2011;6(12):e27837.
[13]Benveniste R E,Morton W R,Clark E A,et al.Inoculation of baboons and macaques with simian immunodeficiency virus/Mne,a primate lentivirus closely related to human immunode⁃ficiency virus type 2[J].J Virol,1988,62(6):2091-2101.
[14]刘存霞,李昌,王茂鹏.新型鸡痘病毒穿梭载体的构建及其体外表达[J].中国兽医科学,2012,42(05):454-460.
[15]Amano H,Morikawa S,Shimizu H,et al.Identification of the canarypox virus thymidine kinase gene and insertion of for⁃eign genes[J].Virology,1999,256(2):280-290.
[16]Lee Huw,Lee Hwa.Application of the polymerase chain reac⁃tion for the diagnosis of fowl poxvirus infection[J].J Virol Methods,1997,63(1-2):113-119.
[17]Scheiflinger F,Falkner F G,Dorner F.Role of the fowlpox vi⁃rus thymidine kinase gene for the growth of FPV recombi⁃nants in cell culture[J].Arch Virol,1997,142(12):2421-2431.
[18]Flynn N M,Forthal D N,Harro C D,et al.Placebo-con⁃trolled phase 3 trial of a recombinant glycoprotein 120 vac⁃cine to prevent HIV-1 infection[J].J Infect Dis,2005,191(5):654-665.
[19]Skinner M A,Laidlaw S M,Eldaghayes I,et al.Fowlpox vi⁃rus as a recombinant vaccine vector for use in mammals and poultry[J].Expert Rev Vaccines,2005,4(1):63-76.
[20]Bertran K,Sá E Silva M,Pantin-Jackwood M J,et al.Protec⁃tion against H7N3 high pathogenicity avian influenza in chick⁃ens immunized with a recombinant fowlpox and an inactivat⁃ed avian influenza vaccines[J].Vaccine,2013,31(35):3572-3576.
[21]Qian C,Chen S,Ding P,et al.The immune response of a re⁃combinantfowlpoxviruscoexpressingtheHA geneofthe H5N1 highly pathogenic avian influenza virus and chicken in⁃terleukin 6 gene in ducks[J].Vaccine,2012,30(44):6279-6286.
[22]Odunsi K,Matsuzaki J,Karbach J,et al.Efficacy of vaccina⁃tion with recombinant vaccinia and fowlpox vectors expressing NY-ESO-1 antigen in ovarian cancer and melanoma patients[J].Proc Natl Acad Sci USA,2012,109(15):5797-5802.
[23]Liska V,Khimani A H,Hofmann-Lehmann R,et al.Viremia and AIDS in rhesus macaques after intramuscular inoculation ofplasmid DNA encodingfull-length SIVmac239[J].AIDS Res Human Retroviruses,1999,15(5):445-450.
[24]Kodama T,Burns D P,Silva D P,et al.Strain-specific neu⁃tralizing determinant in the transmembrane protein of simian immunodeficiency virus[J].J Virol,1991,65(4):2010-2018.
[25]Cannon P M,Matthews S,Clark N,et al.Structure-function studies of the human immunodeficiency virus type 1 matrix protein,p17[J].J Virol,1997,71(5):3474-3483.
[26]Vincent M J,Melsen L R,Martin A S,et al.Intracellular in⁃teraction of simian immunodeficiency virus Gag and Env pro⁃teins[J].J Virol,1999,73(10):8138-8144.
[27]Spies C P,Compans R W.Effects of cytoplasmic domain length on cell surface expression and syncytium-forming ca⁃pacity of the simian immunodeficiency virus envelope glycopro⁃tein[J].Virology,1994,203(1):8-19.
[28]Patterson L J,Daltabuit-Test M,Xiao P,et al.Rapid SIV Env-specific mucosal and serum antibody induction augments cellular immunity in protecting immunized,elite-controller ma⁃caques against high dose heterologous SIV challenge[J].Virolo⁃gy,2011,411(1):87-102.
[29]Kulkarni V,Rosati M,Bear J,et al.Comparison of intrader⁃mal and intramuscular delivery followed in vivo electropora⁃tion of SIV Env DNA in macaques[J].Hum Vaccin Immunoth⁃er,2013,9(10):2081-2094.
[30]Someya K,Xin K Q,Ami Y,et al.Chimeric adenovirus type 5/35 vector encoding SIV gag and HIV env genes affords pro⁃tective immunity against the simian/human immunodeficiency virus in monkeys[J].Virology,2007,367(2):390-397.
猜你喜欢
甘肃畜牧兽医(2022年11期)2023-01-05
华人时刊(2022年9期)2022-09-06
华人时刊(2020年15期)2020-12-14
阅读(科学探秘)(2020年4期)2020-07-04
爱你(2019年21期)2019-06-21
食品科学(2018年10期)2018-05-23
农民致富之友(2017年12期)2018-01-31
广州大学学报(自然科学版)(2015年4期)2015-12-23
西南医科大学学报(2015年1期)2015-08-22
中国当代医药(2015年9期)2015-03-01
