
炭疽芽胞杆菌A16R株FtsE蛋白的可溶性表达和纯化
2014-10-27 09:04李强刘先凯冯尔玲朱力王沛王红杨新华赵美玲姜永强王恒樑
生物技术通讯 2014年1期
李强,刘先凯,冯尔玲,朱力,王沛,王红,杨新华,赵美玲,姜永强,王恒樑
1.白求恩医务士官学校,河北 石家庄 050081;2.军事医学科学院 a.生物工程研究所;b.微生物流行病研究所,北京 100071
炭疽芽胞杆菌(Bacillus anthracis)的芽胞侵入动物或人体可引起包括皮肤炭疽、胃肠道炭疽、肺炭疽在内的3种类型的炭疽疾病,并可被巨噬细胞吞噬而萌发,最终入血导致败血症,是一种非常重要的人畜共患病病原体[1]。炭疽杆菌的芽胞对繁殖体形式不易生存的环境,如干燥、高温、紫外线、电离辐射、极端pH值等的抵抗力极强,提高了菌体感染人或动物的几率;且炭疽杆菌的芽胞是其主要感染形式,并可制备为气溶胶而大面积扩散,成为生物战剂的首选。2001年10月发生的生物恐怖袭击中使用了炭疽芽胞,引起了全世界对于炭疽杆菌芽胞研究的关注[2]。
细菌芽胞形成是菌体分裂的一种特殊形式。我们前期的蛋白质组学研究发现FtsE蛋白在炭疽杆菌芽胞形成过程中明显上调,提示其可能在芽胞形成过程中发挥了重要作用[3]。Garti-Levi等在枯草芽胞杆菌芽胞形成研究中发现,FtsE蛋白不仅可作为ABC转运子摄取外界环境信号,激活Spo0A并起始芽胞形成过程,而且对芽胞形成过程中的不对称分裂起到了关键作用[4]。
为研究炭疽芽胞杆菌中FtsE蛋白的功能,我们利用基因工程技术将其编码序列克隆到原核表达载体,进行了可溶性诱导表达,并优化了亲和层析法大量纯化FtsE蛋白的条件,获得了高纯度目的蛋白,为下一步功能研究奠定了基础。
1 材料和方法
1.1 材料
炭疽杆菌减毒株A16R由军事医学科学院菌种中心提供;大肠杆菌DH5α、BL21(DE3)和表达载体pET-28a为本室保存;Taq DNA聚合酶、限制性内切酶、T4DNA连接酶购自TaKaRa公司;鼠源抗-His抗体购自Pharmacia公司;HRP标记的羊抗鼠IgG购自北京博奥森生物技术有限公司;质粒快速提取试剂盒购自Promega公司;蛋白定量试剂盒购自Amer⁃sham公司;蛋白胨与酵母提取物购自OXOID公司;丙烯酰胺、甲叉双丙烯酰胺购自BBI公司;PEG20000、过硫酸铵、TEMED、EDTA二钠盐、十二烷基磺酸钠(SDS)、考马斯亮蓝G-250、RNase A等购自AMRESCO公司。
低温离心机为SIGMA 3K12;酶联检测仪型号为SS-3000;超声破碎仪为SONICS VC 750型;国产脱色摇床;GeneAmp PCR system为Applied Biosys⁃tems产品;凝胶成像分析仪为TANSILLUMNATOR公司产品;垂直电泳仪购自Amersham公司;蛋白半干电转仪为Bio-Rad公司产品;AKTA FPLC及 Ni-NTA产自Pharmacia公司。
1.2 构建融合表达载体pET28a-ftsE
调取GenBank中炭疽芽胞杆菌ftsE基因序列,利用Primer premier软件,于此基因上下游末端分别插入EcoRⅠ、HindⅢ酶切位点,设计引物(由奥科生物技术公司合成)F(5'-CCGGAATTCATGAAAGCC ATTGCTGGTCT-3')和 R(5'-CCCAAGCTTGCCTTC ATATCCGTATCCTC-3')。
PCR扩增(50 μL反应体系)参数为94℃ 变性10 min,之后以94℃变性30 s、55℃退火30 s、72℃延伸1 min进行30个循环,最后72℃延伸10 min。用凝胶回收试剂盒切胶回收PCR产物,将PCR产物及pET28a质粒分别于37℃水浴条件下用EcoRⅠ、HindⅢ双酶切6 h,用凝胶回收试剂盒回收后在凝胶成像分析仪下定量;根据Novagen pET system Man⁃ual,配制扩增产物与pET载体连接反应体系,16℃反应过夜;将2 μL重组连接反应产物用化学转化法转化到33 μL大肠杆菌DH5α感受态后涂LB(Kan+)平板,分别挑取数个pET-ftsE转化子进行菌落PCR鉴定;对阳性克隆纯培养后提取质粒,双酶切鉴定正确的质粒由奥科公司测序鉴定。
1.3 融合蛋白的诱导表达与Western印迹鉴定
将pET-ftsE重组质粒转化表达宿主菌大肠杆菌BL21(DE3),获得表达外源蛋白的重组大肠杆菌BL21(pET-ftsE),挑取LB(Kan+)平板上的单个重组菌落,接种于5 mL LB(Kan+)液体培养基中,37℃培养10 h,然后取1%接种到新鲜LB(Kan+)液体培养基中,37℃培养140 min后,加入不同浓度的IPTG诱导表达2~4 h,离心收集菌体,煮沸5 min后12 000 r/min离心2 min,取5 μL样品行SDS-PAGE,80 V恒压电泳20 min后,120 V恒压电泳3 h,取凝胶置于染色液中染色30 min,然后用高甲醇脱色液(45%甲醇,47%水,8%冰乙酸)脱色2 h,低甲醇脱色液(5%甲醇,87%水,8%冰乙酸)脱色4 h,观察融合蛋白的表达情况。
取微量融合蛋白与等体积2×SDS缓冲液充分混匀,煮沸5 min,离心后取12 μL上样电泳,将分离的蛋白通过半干电转仪转移到硝酸纤维素膜上,恒压15 V转移1.5 h;用5%脱脂奶粉封闭1 h后,以抗-His抗体为一抗,室温封闭1.5 h,洗膜3次后以HRP标记的羊抗鼠IgG为二抗,室温封闭后吸净PVDF膜表面液体,ECL显色系统显色5 min后用X线胶片在暗室中曝光检测目的蛋白。
1.4 优化融合蛋白的可溶性表达
分别在14、20、30℃条件下用终浓度分别为0.1、0.4、0.5、1.0 mmol/L的IPTG诱导1~4 h,对诱导产物的上清及沉淀分别行SDS-PAGE检测,筛选融合蛋白可溶性表达的条件。
1.5 表达产物的镍柱纯化
按照筛选的诱导条件,将表达融合蛋白的工程菌用IPTG诱导3 h,将2000 mL培养液于4℃条件下8000 r/min离心15 min,收集菌体,重悬于20 mL Ni-NTA柱结合缓冲液中,加入蛋白酶抑制剂,在30%总功率条件下,冰浴超声波破碎20 min(超声2 s,停2 s),4℃、5000 r/min离心30 min,收集上清。
依次用8倍柱体积的双蒸水、4倍柱体积的1 mol/L NaCl、16倍柱体积的双蒸水、8倍柱体积的结合缓冲液缓推过Ni-NTA柱,置4℃冰箱中备用。
处理好的柱子UV约为50,上样3 mL/min至总体积200 mL;用Ni-NTA柱结合缓冲液洗柱3 mL/min,至UV为52;用Ni-NTA柱洗涤缓冲液洗柱3 mL/min,至UV为56;分别用3.3%、10%、20%、30%、50%、70%、100%的洗脱缓冲液洗5个柱体积,收集液体,每管收集1 mL洗脱液,每隔5管取少量洗脱液进行电泳验证,检测纯化蛋白的纯度和蛋白量。
2 结果
2.1 ftsE基因的PCR扩增
GenBank中ftsE基因片段长度为645 bp。由图1可见,扩增出的片段长度与预计相符。
2.2 重组质粒pET28a-ftsE的构建
将表达载体pET28a与ftsE基因的PCR产物分别双酶切后,连接并转化大肠杆菌,挑取转化子提取质粒。将重组质粒pET28a-ftsE用EcoRⅠ、HindⅢ双酶切,结果如图2,1、2、4泳道出现2条电泳带,其中一条分子大的电泳带与载体pET-28a位置相同,另一条分子小的电泳带则位于600 bp左右,与ftsE基因的PCR产物大小相同。
2.3 重组质粒中ftsE基因的核苷酸序列分析
对酶切鉴定正确的菌株传代保存,并挑取单菌落培养10 h后,取1.5 mL菌液送奥科公司测序。测得的ftsE基因的核苷酸序列与GenBank中BA5416序列相同,编码框架完全正确。
2.4 重组蛋白的SDS-PAGE分析及Western印迹
含重组质粒pET-ftsE的大肠杆菌BL21(DE3)经IPTG诱导后,于4℃、5000 r/min离心取菌体,溶解于Ni-NTA柱结合缓冲液后超声波破碎,10 000 r/min离心后分别取上清和包涵体进行SDS-PAGE检测,分析重组FtsE蛋白在大肠杆菌中的表达情况。取微量融合蛋白以抗-His抗体进行Western印迹,发现目的蛋白表达正确(图3)。进一步对表达的温度和诱导的IPTG浓度等条件进行摸索,发现在20℃、0.1 mmol/L IPTG诱导条件下的表达产物主要是可溶性蛋白。
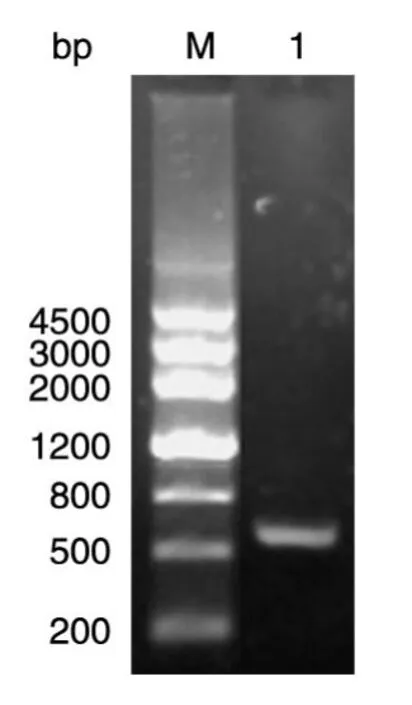
图1 PCR扩增得到ftsE基因

图2 重组质粒的酶切鉴定
2.5 重组FtsE蛋白的分离纯化
经过不断摸索诱导表达条件,在20℃,0.1 mmol/L条件下表达的FtsE主要以可溶性形式存在。将制备得到的全菌体溶液上清用Ni-NTA纯化进行亲和层析,然后用不同浓度的咪唑洗脱缓冲液梯度洗脱得到纯化产物,共得到80管液体,每管1 mL,从中取微量蛋白进行SDS-PAGE,结果见图4。
3 讨论
对大肠杆菌的FtsE蛋白的大量研究表明,其通过参与构建并稳固菌体分裂环及水解细胞壁,在菌体细胞分裂中发挥了重要作用,是菌体分裂相关蛋白[5-6]。对肺炎球菌FtsE蛋白的研究发现,其通过与PCB蛋白的相互作用,参与细胞分裂[7]。对脑膜炎球菌FtsE蛋白的研究发现,其通过与DNA结合,参与摄取外源DNA而进行基因转化[8]。对枯草芽胞杆菌芽胞形成的研究认为,FtsE蛋白作为ABC转运子,参与摄取外源信号物质,起始芽胞形成过程,并在芽胞形成过程中参与细胞的不对称分裂,是芽胞形成中不可或缺的重要蛋白[4]。
炭疽芽胞杆菌中FtsE蛋白的功能还不清楚,因此,我们构建了炭疽芽胞杆菌ftsE基因的原核表达质粒,在大肠杆菌BL21(DE3)中进行了融合表达。因为外源表达的可溶性蛋白具有易分离、活性高、与体内状态更接近的特点,我们摸索了FtsE蛋白的可溶性诱导表达条件,并利用Pharmacia公司的FPLC及Ni-NTA对表达产物进行了纯化,获得了纯度达90%以上的融合蛋白,为下一步抗体制备及其体内外功能的研究奠定了基础。

图3 重组蛋白的诱导表达(左)及Western印迹(右)
图4 纯化洗脱FtsE蛋白的SDS-PAGE
[1]Mock M,Fouet A.Anthrax[J].Annu Rev Microbiol,2001,55:647-671.
[2]Enserink M.Bioterrorism.This time it was real:knowledge of anthrax to the test[J].Science,2001,294(5542):490-491.
[3]Wang Junjun,Ying Tianyi,Wang Hengliang,et al.Two-di⁃mensional reference map of Bacillus anthracis vaccine strain A16R proteins[J].Proteomics,2005,5:4488-4495.
[4]Garti-Levi S,Hazan R,Kain J,et al.The FtsEX ABC trans⁃porterdirects cellulardifferentiation in Bacillussubtilis[J].Mol Microbiol,2008,69(4):1018-1028.
[5]Arends S J,Kustusch R J,Weiss D S.ATP-binding site le⁃sions in FtsE impair cell division[J].J Bacteriol,2009,191(12):3772-3784.
[6]Yang D C,Peters N T,Parzych K R,et al.An ATP-binding cassette transporter-like complex governs cell-wall hydrolysis at the bacterial cytokinetic ring[J].Proc Natl Acad Sci USA,2011,108(45):1052-1060.
[7]Sham L T,Barendt S M,Kopecky K E,et al.Essential PcsB putative peptidoglycan hydrolase interacts with the essential FtsXSpn celldivision protein in Streptococcuspneumoniae D39[J].Proc Natl Acad Sci USA,2011,108(45):1061-1069.
[8]Lang E,Haugen K,Fleckenstein B,et al.Identification of neisserial DNA binding components[J].Microbiology,2009,155:852-862.
猜你喜欢
中国生物防治学报(2022年3期)2022-07-09
当代水产(2022年1期)2022-04-26
微生物学杂志(2021年2期)2021-07-01
小学生必读(中年级版)(2021年12期)2021-03-03
小学生必读(中年级版)(2021年11期)2021-02-22
宁夏医学杂志(2020年4期)2021-01-21
微生物学杂志(2020年2期)2020-12-31
中国调味品(2017年2期)2017-03-20
现代检验医学杂志(2016年3期)2016-11-15
衡阳师范学院学报(2016年3期)2016-07-10
