
载脂蛋白A-I模拟肽D-4F对巨噬细胞源性泡沫细胞清道夫受体A1的抑制作用*
2014-08-13 12:06姚树桐李严严王义围秦树存
中国病理生理杂志 2014年10期
赵 莉, 姚树桐, 陈 军, 苗 成, 李严严, 田 华, 周 健, 翟 雷, 桑 慧, 王义围△, 秦树存△
(1承德医学院附属医院,河北 承德 067000; 泰山医学院 2动脉粥样硬化研究所,山东省高校动脉粥样硬化重点实验室, 3基础医学院,山东 泰安 271000)
作为心脑血管疾病的重要病理基础,动脉粥样硬化(atherosclerosis,AS)严重危害人类的健康,巨噬细胞源性泡沫细胞形成是其重要病理学标志,并在其发生发展中起着重要作用[1]。氧化低密度脂蛋白(oxidizedlow-densitylipoprotein,ox-LDL)是诱导泡沫细胞形成的主要因素。由于巨噬细胞通过清道夫受体A1(scavengerreceptorA1,SR-A1)、CD36等清道夫受体对ox-LDL的摄取是无上限的,导致大量脂质在细胞内蓄积使其变成泡沫细胞。同时由于氧化脂质具有很强的细胞毒效应,最终引起泡沫细胞凋亡、坏死,形成粥样斑块的脂质核心[2]。因此,由清道夫受体介导的对ox-LDL无限制摄取是泡沫细胞形成和AS发生发展的关键,已成为AS治疗的重要靶点。
高密度脂蛋白(high-densitylipoprotein,HDL)具有胆固醇逆向转运、抗炎、抗氧化、保护血管内皮等功能,是公认的具有抗AS作用的“好胆固醇”[3]。但是在AS患者体内HDL水平往往降低,尤其是HDL结构和组成成分的改变使其抗AS作用丧失,甚至表现出促炎、促氧化等特性,成为AS发生发展的危险因素[4]。近年来研究表明HDL主要蛋白成分载脂蛋白A-I(apolipoproteinA-I,ApoA-I)的模拟肽L-4F和D-4F能够促进巨噬细胞中胆固醇流出[5],抑制血脂紊乱所导致的小鼠AS[6],但该模拟肽是否可通过调控清道夫受体表达抑制巨噬细胞内脂质蓄积尚未见报道。本课题组最新研究表明,内质网应激(endoplasmicreticulumstress,ERS)通过上调清道夫受体SR-A1和CD36表达促进巨噬细胞源性泡沫细胞形成[7],故本工作在ox-LDL和ERS诱导剂衣霉素(tunicamycin,TM)诱导的巨噬源性泡沫细胞模型和ERS模型上,研究ApoA-I模拟肽D-4F对SR-A1表达的影响,以探讨其对巨噬源性泡沫细胞形成的调控作用及机制。
材 料 和 方 法
1 材料与试剂
鼠源RAW264.7巨噬细胞购自中国科学院上海生物化学与细胞生物学研究所细胞库;D-4F(Ac-DWFKAFYDKVAEKFKEAF-NH2)和紊乱模拟肽(scrambled D-4F,sD-4F; Ac-DWFAKDYFKKAFVEEFAK-NH2)购自中科亚光多肽服务公司;ox-LDL购自北京协生生物科技有限公司;DMEM高糖培养基和胎牛血清为Gibco产品;RIPA裂解液和BCA蛋白定量试剂盒为Solarbio产品;兔抗SR-A1和葡萄糖调节蛋白78(glucose-regulated protein 78,GRP78)多克隆抗体购自Santa Cruz;4-苯丁酸(4-phenylbutyric acid,PBA)、TM和兔抗β-actin抗体购自Sigma;辣根过氧化物酶标记山羊抗兔IgG为北京中杉金桥公司产品;增强化学发光(enhanced chemiluminescence,ECL)试剂盒和PVDF膜分别为Pierce和Millpore产品;Trizol试剂为Invitrogen产品;cDNA合成试剂盒和Real Master Mix(SYBR Green)试剂盒购自北京天根公司;组织/细胞总胆固醇(total cholesterol,TC)测定试剂盒和DiI-ox-LDL分别购自北京普利莱公司和北京协生生物公司;其余试剂均为分析纯产品。
2 方法
2.1细胞培养与处理 用高糖DMEM培养基(含10%胎牛血清、青霉素1×105U/L和链霉素100 mg/L)于37 ℃、5% CO2培养箱中培养RAW264.7巨噬细胞。处理前换含1%血清DMEM培养基培养12 h,然后给予不同浓度的D-4F(12.5、25和50 mg/L)、sD-4F(50 mmol/L)处理1 h或者5 mmol/L PBA处理30 min后,再加入ox-LDL(100 mg/L)继续培养12 h。另外培养巨噬细胞给予50 mg/L D-4F 或sD-4F 处理1 h,再加入2 mg/L TM处理4 h。
2.2MTT法检测细胞活性 将细胞以1×104/well接种于96孔培养板内,细胞经处理后,加入MTT(0.5g/L),37 ℃继续培养 4h。弃上清后每孔加 200μL二甲亚砜(dimethylsulfoxide,DMSO),振荡摇匀,用InfiniteF200型多功能酶标仪(Tecan)在490nm处测定各孔吸光度(absorbance,A)。以对照组细胞活力为100%,其余各组细胞活力以其A值占对照组A值的百分比表示。
2.3试剂盒检测细胞内TC的含量 按照组织/细胞TC测定试剂盒说明书测定细胞内TC,并以细胞每克总蛋白中所含的TC量(μmol/g protein)来表示。
2.4Westernblotting检测SR-A1和GRP78蛋白的表达 细胞经处理后,按照RIPA蛋白提取试剂盒说明书提取各组细胞蛋白,经定量、变性处理并进行SDS-PAGE(10%分离胶)分离后电转印至PVDF膜,5%脱脂奶粉封闭后分别用兔抗SR-A1 (1∶300)、GRP78 (1∶400)和β-actin抗体(1∶800)室温孵育3h,洗膜后用辣根过氧化物酶标记相应Ⅱ抗孵育2h。免疫条带用ECL法显示,暗室曝光,采用Image-ProPlus软件分析蛋白条带累积吸光度(integratedabsorbance,IA)值,以靶蛋白IA值/β-actinIA值的比值反映靶蛋白相对水平。
2.5实时荧光定量聚合酶链反应(real-time PCR)技术检测SR-A1 mRNA的表达 按照Trizol试剂说明提取各组细胞总RNA。取RNA样品1 μg,按RNA逆转录试剂盒说明书合成cDNA产物。按荧光实时定量PCR试剂盒说明书准备20 μL PCR扩增反应体系:SYBR Green 9 μL,上、下游引物各2 μL,cDNA 2 μL,双蒸水5 μL。在荧光定量PCR仪(Rotor-Gene Q型,Qiagen)上反应,条件为94 ℃预变性10 min,94 ℃ 20 s,55 ℃ 30 s,68 ℃ 30 s,40个循环。β-actin为内参照,利用2-ΔΔCt公式计算目的基因mRNA的表达情况。SR-A1上游引物5’-GTTAAAGGTGATCGGGGACA-3’,下游引物5’-TCCCCTTCTCTCCCTTTTGT-3’,扩增产物长度118 bp;β-actin上游引物5’-CGGGGACCTGACTGACTACC-3’,下游引物5’- AGGAAGGCTGGAAGAGTGC-3’,扩增产物长度251 bp,由上海生工公司合成。
2.6DiI-ox-LDL摄取的分析 参照既往报道方法[7]操作,即给予巨噬细胞50mg/LD-4F或sD-4F处理1h,再加入2mg/LTM处理4h后,加入DiI-ox-LDL(50mg/L)继续培养4h,避光条件下以PBS润洗3次,并以RIPA裂解液裂解细胞,BCA法蛋白定量。2 000r/min离心5min,将上清200μL加入96孔板,用多功能酶标仪(InfiniteF200型,Tecan)检测荧光强度(激发波长530nm,发射波长590nm)。将DiI-ox-LDL稀释成0、5、10、15、20、25、50和100mg/L,相同条件下检测荧光强度,制作标准曲线。ox-LDL摄取量以mg/gprotein表示。
3 统计学处理
数据用均数±标准差(mean±SD)表示,采用SPSS 13.0 for Windows 统计学软件进行单因素方差分析,组间均数比较用SNK法,以P<0.05为差异有统计学意义。
结 果
1 D-4F减轻ox-LDL所诱导的巨噬细胞损伤
ox-LDL显著诱导RAW264.7巨噬细胞损伤,其细胞活性较正常对照组降低43.7%(P<0.01);与PBA相似,D-4F可减轻ox-LDL所诱导的细胞损伤,细胞活性明显增加且呈浓度依赖性(P<0.05或P<0.01);ox-LDL+sD-4F组与ox-LDL组比较细胞活性无显著差异(P>0.05),见图1。

Figure 1. D-4F (mg/L) inhibited ox-LDL-induced macrophage injury.Mean±SD.n=6.*P<0.05,**P<0.01 vs control group;#P<0.05, ##P<0.01 vs ox-LDL group.
2 D-4F减轻ox-LDL所诱导的RAW264.7细胞内胆固醇的蓄积
细胞内TC测定结果显示,与正常对照组比较,ox-LDL组巨噬细胞内TC含量显著增加(P<0.01),而D-4F(12.5、25和50 mg/L)明显抑制ox-LDL所诱导的细胞内胆固醇蓄积,其TC含量分别为ox-LDL 组的69.9%、57.6%和53.4%(P<0.05或P<0.01);PBA也可明显抑制ox-LDL所诱导的细胞内胆固醇蓄积(P<0.05),见图2。
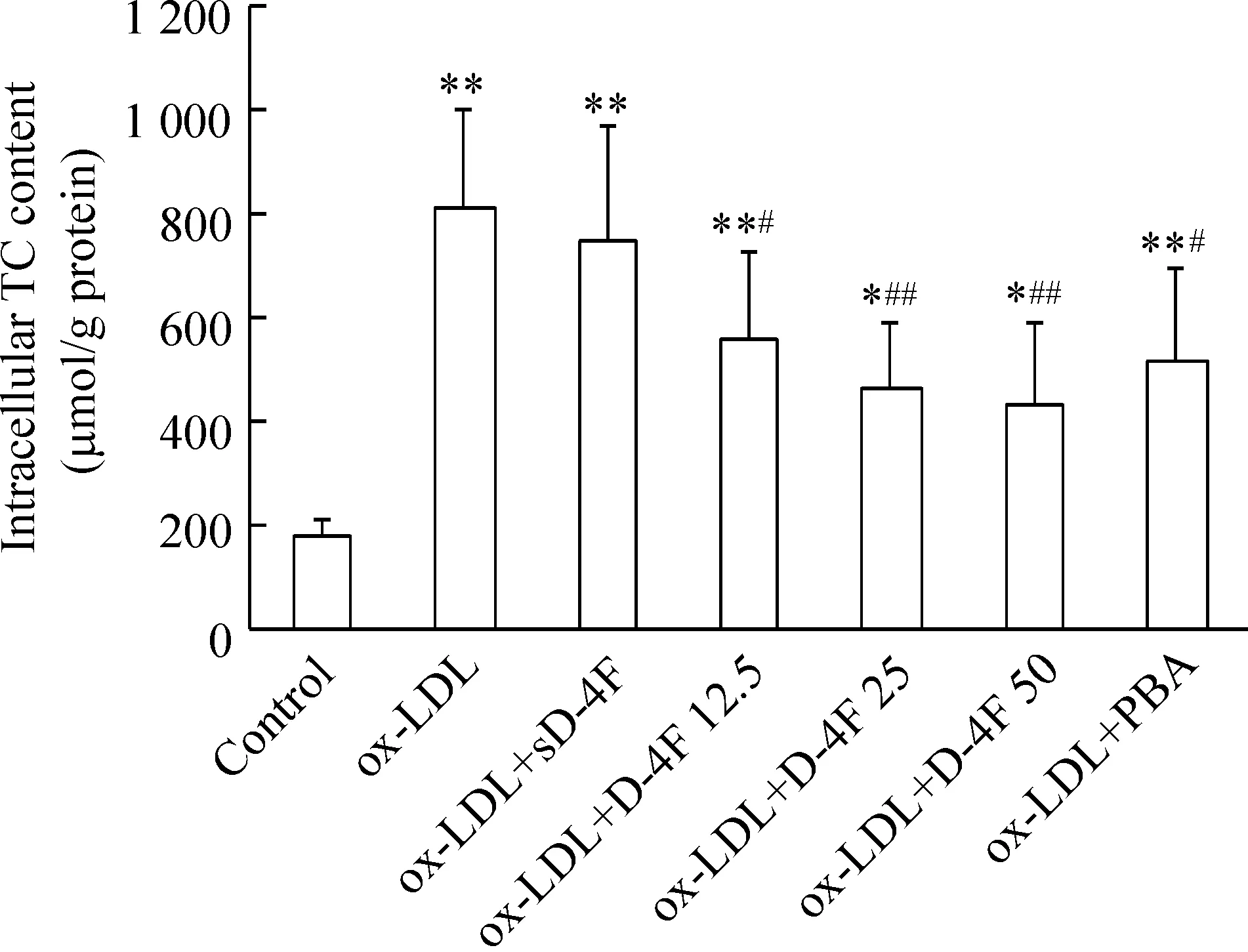
Figure 2. D-4F (mg/L) reduced ox-LDL-induced intracellular cholesterol accumulation in RAW264.7 cells. Mean±SD. n=6.*P<0.05, **P<0.01 vs control group; #P<0.05, ##P<0.01 vs ox-LDL group.
3 D-4F对ox-LDL所诱导的SR-A1和GRP78蛋白表达的影响
与对照组比较,ox-LDL处理组细胞SR-A1和GRP78蛋白水平明显上调(P<0.01),而D-4F同PBA相似,显著抑制ox-LDL所诱导的SR-A1和GRP78蛋白上调,并且呈浓度依赖性(P<0.05),见图3。

Figure 3. The effect of D-4F (mg/L) on ox-LDL-induced SR-A1 and GRP78 protein expression in RAW264.7 cells.Mean±SD.n=3.*P<0.05, **P<0.01 vs control group; #P<0.05 vs ox-LDL group.
4 D-4F对ox-LDL所诱导的SR-A1 mRNA表达的影响
与对照组比较,ox-LDL明显上调SR-A1 mRNA水平(P<0.01),而D-4F显著抑制ox-LDL所诱导的SR-A1 mRNA上调,且呈浓度依赖性(P<0.05或P<0.01),见图4。
5 D-4F对TM所诱导的SR-A1和GRP78表达的影响
如图5所示,与对照组比较,TM可明显上调巨噬细胞SR-A1和GRP78蛋白水平(P<0.01),而D-4F显著抑制TM所诱导的SR-A1和GRP78蛋白表达,分别下降45.5%和42.1%(P<0.05)。
6 D-4F对TM所诱导的巨噬细胞ox-LDL摄取的影响
Dil-ox-LDL吞噬实验结果(图6)显示,与对照组比较,TM明显促进巨噬细胞对Dil-ox-LDL的摄取(P<0.01),而D-4F对TM所诱导的巨噬细胞ox-LDL摄取有明显抑制作用,为TM组的62.9%(P<0.05)。

Figure 4. The effect of D-4F (mg/L) on ox-LDL-induced mRNA expression of SR-A1 in RAW264.7 cells. Mean±SD.n=4.*P<0.05, **P<0.01 vs control group; #P<0.05, ##P<0.01 vs ox-LDL group.

Figure 5. The effect of D-4F on tunicamycin (TM)-induced SR-A1 and GRP78 expression in RAW264.7 cells.Mean±SD.n=3.*P<0.05, **P<0.01 vs control group; #P<0.05 vs TM group.

Figure 6. The effect of D-4F on tunicamycin (TM)-induced uptake of ox-LDL in RAW264.7 cells.Mean±SD.n=4. *P<0.05, **P<0.01 vs control group; #P<0.05 vs TM group.
讨 论
HDL通过逆转运胆固醇、抗脂质氧化、抗炎、抗血栓形成、保护内皮细胞等功能发挥其抗AS作用,但是近年研究发现HDL含量与其功能不成比例,在AS患者体内HDL亚组分改变,如蛋白成分氧化修饰或糖基化修饰后其抗AS功能明显降低,甚至表现出促炎、促氧化等促AS特性[4, 8]。ApoA-I是HDL的主要蛋白成分和功能执行者,但由于其分子量大,制造起来非常困难并且昂贵。近年来运用生物技术开发研制的具有18个氨基酸肽链片段的ApoA-I模拟肽L-4F和D-4F能够促进巨噬细胞中胆固醇流出、增加前β-HDL的形成、减少脂蛋白氧化,使失功能HDL变成功能HDL,从而发挥抗AS功能[5-6,8]。本实验结果显示, D-4F可以显著抑制ox-LDL所诱导的巨噬细胞细胞损伤,并减轻细胞内胆固醇蓄积。
由清道夫受体介导的对ox-LDL无限制摄取是泡沫细胞形成和AS发生发展的关键。SR-A1是巨噬细胞表面的糖蛋白,特异性识别并且结合ox-LDL,且其介导的巨噬细胞对ox-LDL的摄取不受细胞内胆固醇水平的负反馈调节,导致细胞内脂质无限制蓄积,形成AS粥样斑块特征性标志——泡沫细胞,进而发生凋亡、坏死,导致易损斑块形成,成为急性心血管事件的重要病理基础[2,9]。利用siRNA干扰技术沉默SR-A表达可减少AS斑块及泡沫细胞的形成,提示抑制SR-A介导的巨噬源性泡沫细胞形成可能是AS防治新的突破口[10]。本工作结果显示,D-4F在转录和蛋白水平均明显抑制ox-LDL所诱导的SR-A1上调,且呈浓度依赖性,提示D-4F对巨噬细胞泡沫化和损伤的抑制作用与下调SR-A1表达有关。
近年来ERS在AS发生发展中的作用已受到广泛关注,证实ERS在脂质代谢异常、炎症反应激活、细胞凋亡等过程中起着重要作用,同时高同型半胱氨酸、高血糖等独立的心血管危险因子也被证明可能通过诱导ERS而诱发AS[11]。研究报道软脂酸酯可上调巨噬细胞ox-LDL受体1(ox-LDL receptor 1,LOX-1)表达,而沉默ERS关键信号分子肌醇需求激酶1(inositol-requiring enzyme 1,IRE1)和双链RNA依赖的蛋白激酶R样ER激酶(PKR-like ER kinase,PERK)则明显拮抗软脂酸酯对LOX-1的诱导作用[12]。本课题组最近已证实沉默ERS另一关键信号分子活化转录因子6(activating transcription factor 6,ATF6)及其下游分子伴侣GRP78后可减轻ox-LDL所诱导的巨噬细胞内脂质蓄积[7, 13],且ERS抑制剂PBA可抑制ox-LDL所诱导的巨噬细胞SR-A1上调,提示ERS通过上调清道夫受体表达介导ox-LDL所诱导的巨噬细胞泡沫化过程。本实验结果显示,与PBA相似,D-4F不仅可减轻ox-LDL所诱导的巨噬细胞损伤和细胞内胆固醇蓄积,而且抑制SR-A1和GRP78蛋白上调。本课题组和文献既往报道ERS诱导剂毒胡萝卜素和TM可上调巨噬细胞清道夫受体CD36、SR-A1和LOX-1表达[7,12,14]的结果一致。为了进一步证实D-4F是否通过调控ERS信号途径减轻ox-LDL 所诱导的SR-A1上调,我们在TM所诱导的巨噬细胞ERS模型上研究了D-4F对SR-A1表达和ox-LDL摄取的影响,结果显示,D-4F可显著抑制TM所诱导的巨噬细胞SR-A1和GRP78上调及对ox-LDL的吞噬,提示D-4F可能通过抑制ERS信号途径减轻 SR-A1所介导的巨噬细胞对ox-LDL的摄取。
综上所述,本研究通过细胞实验发现ApoA-I模拟肽D-4F可以通过抑制GRP78介导的ERS信号途径减轻ox-LDL所诱导的清道夫受体SR-A1表达,从而减轻巨噬细胞内胆固醇蓄积和细胞损伤,发挥抗AS功能。CD36是巨噬细胞膜上介导ox-LDL摄取的另一重要清道夫受体,而ATP结合盒转运蛋白A1(ATP-binding cassette transporter A1,ABCA1)和ATP结合盒转运体G1(ATP-binding cassette transporter G1,ABCG1)是介导细胞内胆固醇流出的重要载体,D-4F是否也可通过调控上述受体表达进而影响到巨噬细胞内脂质的蓄积正在进一步研究中。
[参 考 文 献]
[1] Badimon L, Storey RF, Vilahur G. Update on lipids, inflammation and atherothrombosis[J]. Thromb Haemost, 2011, 105(Suppl 1):S34-S42.
[2]SeimonT,TabasI.Mechanismsandconsequencesofmacrophageapoptosisinatherosclerosis[J].JLipidRes, 2009, 50(Suppl):S382-S387.
[3] Farmer JA, Liao J. Evolving concepts of the role of high-density lipoprotein in protection from atherosclerosis[J]. Curr Atheroscler Rep, 2011, 13(2):107-114.
[4]TothPP,DavidsonMH.High-densitylipoproteins:markerofcardiovascularriskandtherapeutictarget[J].JClinLipidol, 2010, 4(5): 359-364.
[5] Xie Q, Zhao SP, Li F. D-4F, an apolipoprotein A-I mimetic peptide, promotes cholesterol efflux from macrophages via ATP-binding cassette transporter A1[J]. Tohoku J Exp Med, 2010, 220(3):223-228.
[6]QinS,KamannaVS,LaiJH,etal.ReverseD-4F,anapolipoprotein-AImimeticpeptide,inhibitsatherosclerosisinApoE-nullmice[J].JCardiovascPharmacolTher, 2012, 17(3): 334-343.
[7] Yao S, Miao C, Tian H, et al. Endoplasmic reticulum stress promotes macrophage-derived foam cell formation by up-regulating cluster of differentiation 36 (CD36) expression[J]. J Biol Chem, 2014, 289(7):4032-4042.
[8]ImaizumiS,NavabM,MorgantiniC,etal.Dysfunctionalhigh-densitylipoproteinandthepotentialofapolipoproteinA-Imimeticpeptidestonormalizethecompositionandfunctionoflipoproteins[J].CircJ, 2011, 75(7):1533-1538.
[9] Zhou F, Pan Y, Huang Z, et al.Visfatin induces cholesterol accumulation in macrophages through up-regulation of scavenger receptor-A and CD36[J]. Cell Stress Chaperones, 2013, 18(5):643-652.
[10]MakinenPI,LappalainenJP,HeinonenSE,etal.SilencingofeitherSR-AorCD36reducesatherosclerosisinhyperlipidaemicmiceandrevealsreciprocalupregulationofthesereceptors[J].CardiovascRes, 2010, 88(3):530-538.
[11] 姚树桐, 秦树存. 内质网应激在动脉粥样硬化发生、发展和防治中的作用[J]. 中国病理生理杂志, 2014, 30(2):364-368.
[12]IshiyamaJ,TaguchiR,AkasakaY,etal.UnsaturatedFAspreventpalmitate-inducedLOX-1inductionviainhibitionofERstressinmacrophages[J].JLipidRes, 2011, 52(2):299-307.
[13] Yao S, Zong C, Zhang Y, et al. Activating transcription factor 6 mediates oxidized LDL-induced cholesterol accumulation and apoptosis in macrophages by up-regulating CHOP expression[J]. J Atheroscler Thromb, 2013, 20(1):94-107.
[14]HuaY,KandadiMR,ZhuM,etal.Tauroursodeoxycholicacidattenuateslipidaccumulationinendoplasmicreticulum-stressedmacrophages[J].JCardiovascPharmacol, 2010, 55(1):49-55.
猜你喜欢
青少年科技博览(中学版)(2022年11期)2023-01-07
作文周刊·小学一年级版(2022年20期)2022-05-07
趣味(数学)(2021年4期)2021-08-05
小天使·一年级语数英综合(2019年9期)2019-11-10
读者(2018年23期)2018-11-20
农家科技下旬刊(2018年5期)2018-07-29
故事作文·低年级(2018年2期)2018-02-22
中成药(2017年7期)2017-11-22
老年世界(2017年5期)2017-06-13
农家科技下旬刊(2016年4期)2016-11-19
