
超抗原葡萄球菌肠毒素A拮抗伊马替尼抑制T细胞活化作用的研究*
2014-05-16 02:17颜宇辉陈小华王冠明李扬秋
中国病理生理杂志 2014年3期
颜宇辉, 陈小华, 王冠明, 林 晨△, 李扬秋
(暨南大学1医学院微生物与免疫学系,2血液研究所,广东广州 510632)
超抗原葡萄球菌肠毒素A拮抗伊马替尼抑制T细胞活化作用的研究*
颜宇辉1, 陈小华1, 王冠明1, 林 晨1△, 李扬秋2
(暨南大学1医学院微生物与免疫学系,2血液研究所,广东广州 510632)
目的:探讨葡萄球菌肠毒素A(staphylococcal enterotoxin A,SEA)对甲磺酸伊马替尼(imatinib mesylate,IM)抑制T淋巴细胞活化的影响。方法:2 mg/L SEA和50 nmol/L IM联合作用于Jurkat细胞24 h后,用实时荧光定量PCR技术检测CD3ε和ζ链mRNA表达水平变化;Western blotting技术检测CD3ε和ζ链蛋白水平变化。结果:单纯IM组CD3ε和ζ链mRNA及蛋白表达均表现下调;SEA与IM联合用药组可以逆转IM下调CD3ε和ζ链mRNA及蛋白表达水平作用。SEA拮抗IM抑制作用中,对CD3ε链mRNA表达上调的作用明显大于CD3ζ链。结论:SEA能拮抗IM对T细胞CD3ε和ζ链表达的抑制作用。
超抗原;葡萄球菌肠毒素A;伊马替尼;T淋巴细胞;抗原,CD3
材料和方法
1 材料
金黄色葡萄球菌肠毒素A(Sigma);伊马替尼(Selleck Chemicals);Cell Counting Kit-8 CCK-8;(日本同仁化学研究所);胎牛血清(杭州四季青生物工程材料有限公司);Trizol试剂(北京康为世纪生物有限公司)。逆转录试剂盒和Real Master Mix试剂盒(日本东洋纺)。Jurkat细胞来源于暨南大学血液病研究所。
2 方法
2.1 IM对Jurkat细胞浓度的选择 培养体系如下: Jurkat细胞1.0×109/L,含10%灭活胎牛血清的RPMI-1640培养液,1×105U/L青霉素,100 mg/L链霉素。各孔分别加入50、100、200和300 nmol/L IM,对照孔加RPMI-1640培养液,设置空白孔,每孔做3复孔,放置于37℃、5%CO2培养箱内培养24 h。培养结束时,每个实验孔内加入10 μL CCK-8试剂后,再继续培养4 h,用双波长测定吸光度(A),检测波长为450 nm,参比波长为600 nm。各组抑制率=1-(实验孔A值-空白孔A值)/(对照孔A值-空白孔A值)。
2.2 SEA联合IM与Jurkat细胞培养 分4组:2 mg/L SEA组、50 nmol/L IM组、SEA+IM联合组和对照组。将状态良好的 Jurkat细胞调节为浓度为1.0×109/L,接种于12孔板中,分别加入相应浓度的SEA与IM,总体积1.5 mL,37℃、5%CO2培养箱中培养24 h。
2.3 实时荧光定量PCR 采用Trizol试剂处理样品细胞,氯仿、异丙醇常规提取RNA,合成cDNA链,利用SYBR Green I染料检测各样本的CD3ε和ζ链mRNA表达情况,并将β2M作为内参照,采用相对定量法分析各组CD3ε和ζ链mRNA表达水平的差异[3]。根据文献[5]设计CD3ζ链序列引物 (ACCESSION No.J04132)。CD3ζ链上游引物5'-GCC AGA ACC AGC TCT ATA AC-3',下游引物5'-TAG GCC TCC GCC ATC TTA TC-3',扩增目的片段长度166 bp。根据CD3ε链的序列设计引物(ACCESSION No.NM_000f733)[3]。扩增 CD3ε链上游引物5'-TCC CAA CCC AGA CTA TGA GC-3',下游引物5'-CAA GAC TAG CCC AGG AAA CAG-3',扩增目的片段的长度为158 bp。β2M上游引物5'-TAC ACT GAA TTC ACC CCC AC-3',下游引物5'-CAT CCA ATC CAA ATG CGG CA-3',扩增目的片段长度145 bp。
总反应体积为20 μL,包括Real Master Mix 10 μL,0.5 mmol/L上、下游引物,1 μL cDNA,8 μL蒸馏水。在95℃、1 min预变性后,共进行40个循环扩增,每一循环包括95℃ 30 s,60℃ 30 s,72℃ 15 s。并在72℃读板1次。随后,以0.15℃/s变化速度从65℃到95℃每隔1 s记录1次荧光值,获得熔解曲线。反应在MJ Research DNA Engine Opticon 2荧光定量PCR仪(Bio-Rad)中进行。
以β2M为内参照,利用Ct值计算各组CD3ε链和ζ链表达的相对量。并用相对定量公式:2-ΔΔCt计算诱导组与空白对照组CD3ε链和ζ链的表达差异,其中ΔΔCt=ΔCt(诱导组)-ΔCt(对照组),ΔCt=Ct (ζ/ε)-Ct(β2M),而基因差异表达的倍数即为2-ΔΔCt。
清水塘22号这座普通的平房,既是中共湘区委员会的机关所在地,又是毛泽东与杨开慧于1920年冬结成革命伴侶后所组建的第一个真正意义上的家。
2.4 Western blotting Jurkat细胞分4组:2 mg/L SEA组、50 nmol/L IM组、SEA+IM联合组和对照组。24 h后收集细胞,加60 μL细胞裂解液,混匀,冰上裂解30 min,每隔10 min振荡1次,4℃12 000 ×g离心10 min,取上清,与5×上样液4∶1混合,沸水浴10 min,-20℃保存。BCA法测定样品蛋白浓度,等量蛋白30 μg 10%SDS-PAGE,用半干转方式将蛋白转移到PVDF膜上,将膜在含5%脱脂奶粉TBST中室温封闭1 h,TBST洗3次,每次5 min。放入Ⅰ抗4℃过夜,TBST洗3次,每次5 min,将膜放入Ⅱ抗,室温孵育1 h,TBST洗3次,每次5 min,ECL显色。目的条带用ImageJ分析软件进行灰度值分析,以GAPDH内参照进行校正。
3 统计学处理
利用SPSS 16.0软件对数据分析。数据用均数±标准差(mean±SD)表示,多组间比较用单因素方差分析,两两比较采用LSD-t检验,以P<0.05为差异有统计学意义。
结果
1 IM对Jurkat细胞增殖影响
不同浓度IM作用于Jurkat细胞24 h,CCK-8测定其对细胞增殖抑制作用的影响。结果显示随着IM浓度的增加,IM对Jurkat细胞的抑制逐渐增加,见图1。
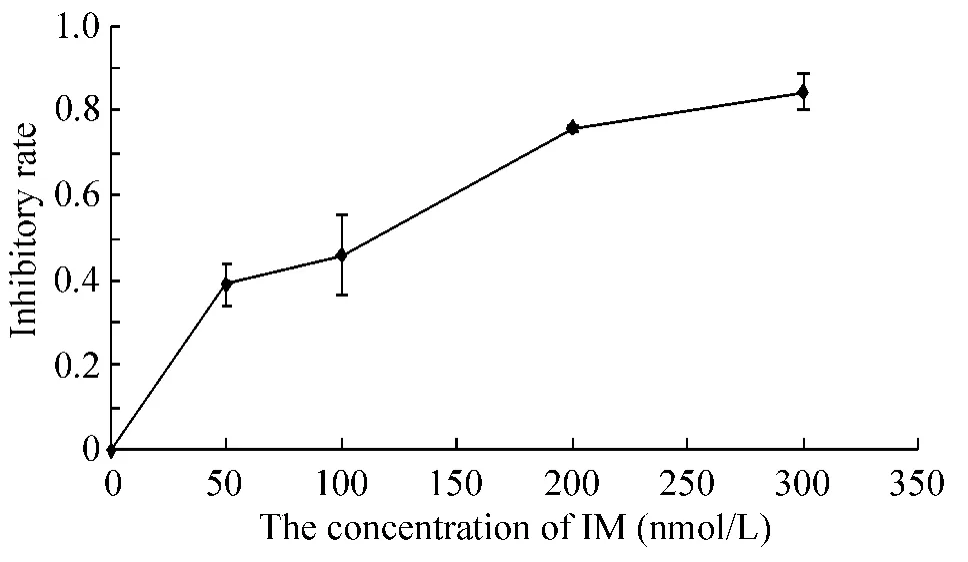
Figure 1.The effects of different concentrations of IM on the proliferation of Jurkat cells after 24 h.Mean±SD.n=3.图1 不同浓度IM作用Jurkat细胞24 h对其增殖的影响
2 PCR产物鉴定
β2M、CD3ε和CD3ζ熔解曲线均只有单一峰,特异性良好,见图2。
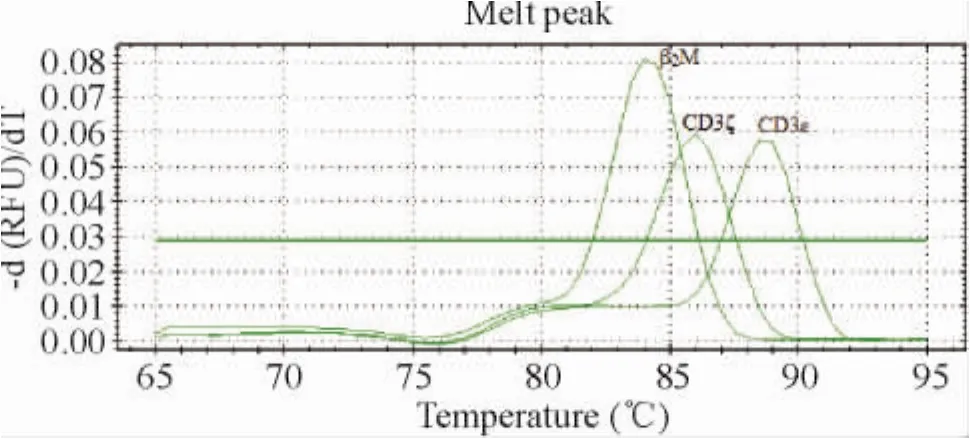
Figure 2. Real-time fluorescence quantitative PCR melting curves of β2M,CD3ε and CD3ζ。图2 实时荧光定量PCR测定β2M、CD3ε和CD3ζ的熔解曲线
3 SEA、IM及SEA+IM对Jurkat细胞CD 3ε和ζ链mRNA表达的影响
根据实时定量PCR的相对定量公式计算基因表达异常倍数值,2-ΔΔCt>1表明表达量增高,2-ΔΔCt<1表明表达量降低。单纯SEA(2 mg/L)作用于Jurkat细胞,CD3ε和ζ链mRNA表达水平明显上调;单纯IM(50 nmol/L)作用,CD3ε和CD3ζ链mRNA表达水平明显下调;而在SEA+IM作用下,与同组IM组比较,被抑制的CD3ε和ζ链mRNA表达水平得以逆转(P<0.05);并且,SEA+IM对CD3ε链mRNA表达影响明显强于对CD3ζ链mRNA表达水平影响(P<0.05),见图3。
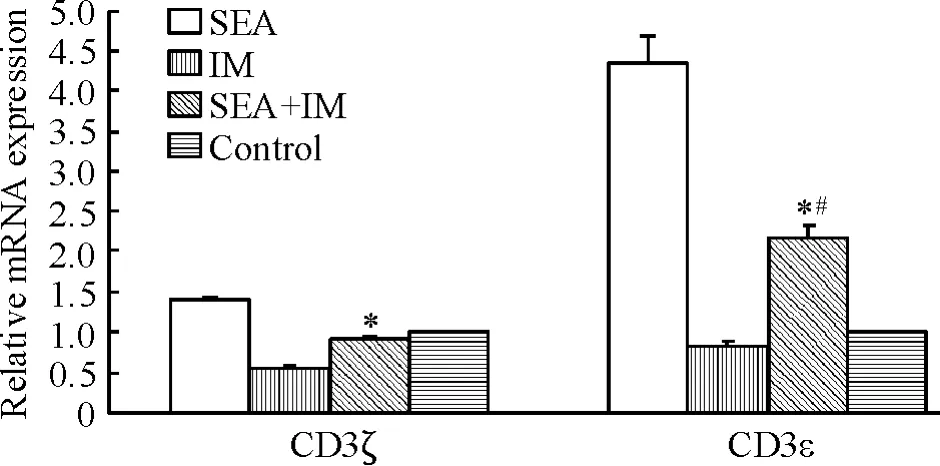
Figure 3.Effects of SEA,IM and SEA+IM on the mRNA expression of CD3ε and CD3ζ in Jurkat cells.Mean± SD.n=3.*P<0.05 vs IM group;#P<0.05 vs CD3ζ in the same group.图3 SEA、IM及SEA+IM对Jurkat细胞CD3ε链与CD3ζ链mRNA表达的影响
4 SEA、IM及SEA+IM对Jurkat细胞CD3ε和ζ链蛋白表达的影响
单独SEA组CD3ε链蛋白表达上调(P<0.05); CD3ζ链变化不大;单独IM组CD3ε和ζ链蛋白表达均下调(P<0.05)。与单纯IM组相比,SEA+IM组逆转IM下调CD3ε和ζ链蛋白的表达(P<0.05),结果类似于mRNA表达水平,见图4。
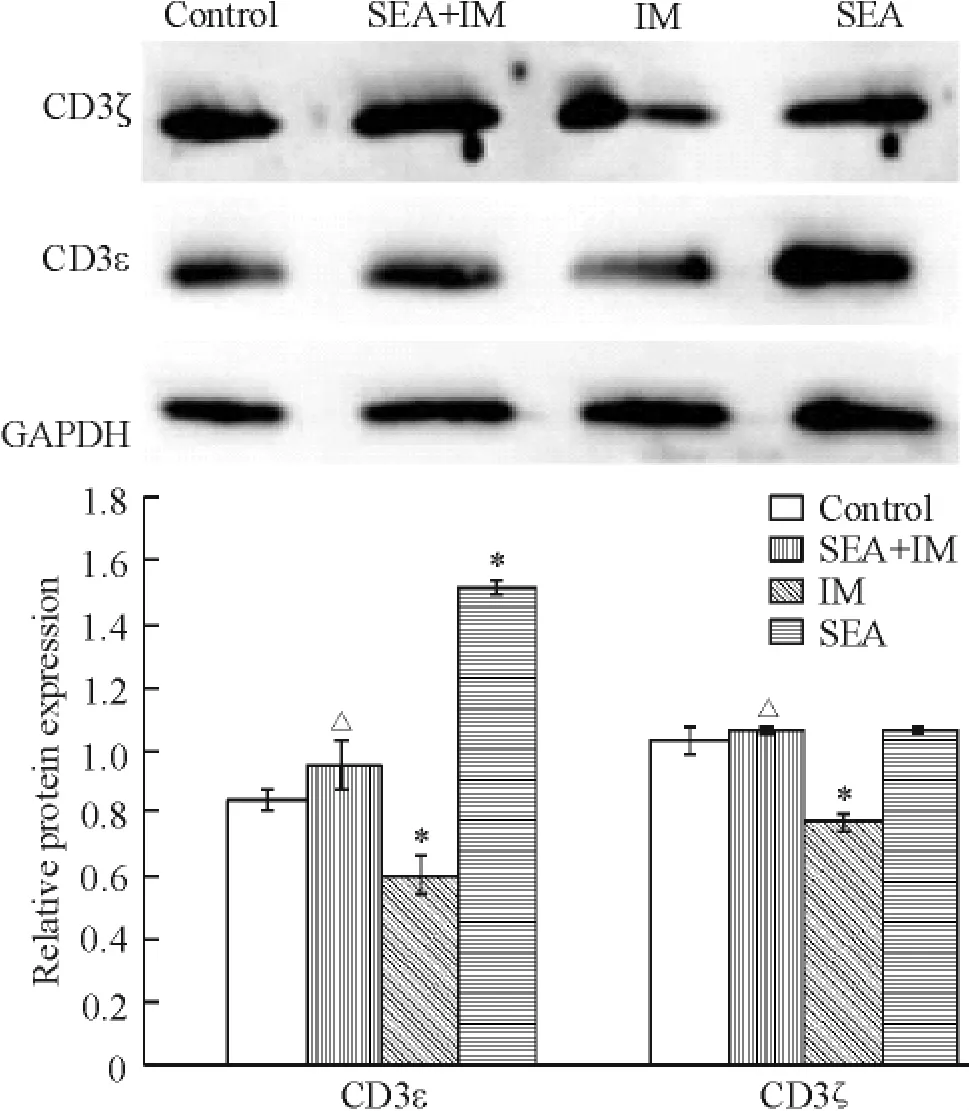
Figure 4.The effects of SEA,IM and SEA+IM on protein expression of CD3ε and ζ chains measured by Western blotting.Mean±SD.n=3.*P<0.05 vs control group;△P<0.05 vs IM group.图4 Western blotting测定SEA、IM及SEA+IM对Jurkat细胞CD3ε链与CD3ζ链蛋白表达的影响
讨论
IM作为一线药物治疗CML获得巨大成功,明显改善了疾病的生存,但仍存在耐药和复发,除了IM诱导性突变之外,病人免疫功能缺陷,缺乏清除残留病能力,也可能是其中一个协同因素。研究表明,IM及随后开发研制的新型靶向药物(克服耐药问题),均具有抑制T细胞的免疫应答作用[1]。如何增强特异性T细胞免疫应答,彻底清除白血病干细胞是十分需要重视并加以研究的问题。
TCR/CD3复合体由TCR受体与CD3分子通过盐桥连接而成,CD3分子在TCR信号传递中起关键作用,CD3分子由γ、δ、ε、ζ和η 5条肽链构成,分别形成CD3γε、CD3δε和CD3ζζ二聚体,其胞浆区含有免疫受体酪氨酸活化基序(immunoreceptor tyrosinebased activation motif,ITAM),负责介导T细胞激活信号转导[2-7]。CD3ε和ζ链表达水平下调均可影响T细胞活化水平,与临床肿瘤[8-9]、自身免疫性疾病[10]及慢性感染[11]等疾病发生相关。
目前,已有不少关于超抗原抗肿瘤研究的报道[12-13],通过超抗原针对性地激活T淋巴细胞靶向杀伤肿瘤细胞,以达到治疗肿瘤的目的。我们也报道了SEA具有增强抗原诱导特异性T细胞杀伤急性粒细胞白血病和CML的作用[14-16]。据此,我们推测SEA具有拮抗IM导致T细胞抑制的可能性。Jurkat细胞株是一商品化的传代T细胞系,本实验采用SEA与IM共同作用Jurkat细胞,利用实时荧光定量PCR检测CD3ε和ζ链mRNA表达;并采用Western blotting检测这2种蛋白表达。结果表明,单纯SEA组T细胞CD3ε和ζ链mRNA表达水平升高,与之前的研究结果一致[3-4]。单纯IM组T细胞CD3ε和ζ链mRNA表达水平降低,也证实IM通过抑制T细胞活化信号通路中上游信号分子的表达水平,导致T细胞活化受阻[17]。而在联合组中,SEA+IM共同作用于T细胞后,表现出逆转IM下调T细胞CD3ε和ζ链mRNA与蛋白表达水平的作用。本研究结果也显示,各组在mRNA水平上CD3ε链表达受到的影响比CD3ζ链明显,具体原因有待进一步研究。TCR信号通过ITAM的激活转导。TCR复合体中的ITAM分布在CD3ζζ和CD3γε/δε多肽链中。虽然CD3ζ具有3个ITAM,CD3ε链有1个ITAM。但有研究表明:即使在不存在TCRζζ的情况下,CD3γε/δε依然可以激活T细胞[18],提示CD3γε/δε在T细胞激活过程中也具有重要的激活作用。
综上所述,超抗原 SEA能拮抗 IM对 T细胞CD3ε和ζ链的抑制作用,为逆转IM临床治疗CML导致的免疫抑制提供了依据。为通过激活特异性T细胞进一步清除CML微小残留病变提供新的思路与治疗手段。
[1]Blake SJ,Hughes TP,Lyons AB.Drug-interaction studies evaluating T-cell proliferation reveal distinct activity of dasatinib and imatinib in combination with cyclosporine A[J].Exp Hematol,2012,40(8):612-621.
[2]陈小华,颜宇辉,王冠明,等.伊马替尼抑制Jurkat T细胞增殖与影响A20和NF-κB表达相关[J].免疫学杂志,2013,29(10):854-858.
[3]田红霞,高永鹏,林 晨,等.超抗原SEA对K562细胞诱导脐带血单个核细胞上CD3ε链表达的影响[J].细胞与分子免疫学杂志,2010,26(12):1175-1177.
[4]高永鹏,林 晨,田红霞,等.SEA联合K562细胞体外诱导正常人脐带血单核细胞TCRζ链表达的作用[J].暨南大学学报:自然科学与医学版,2010,31(2): 154-157.
[5]Sun ZY,Kim ST,Kim IC,et al.Solution structure of the CD3εδ ectodomain and comparison with CD3εγ as a basis for modeling T cell receptor topology and signaling[J].Proc Natl Acad Sci U S A 2004,101(48):16867-16872.
[6]Wang Y,Becker D,Vass T,et al.A conserved CXXC motif in CD3ε is critical for T cell development and TCR signaling[J].PLoS Biol,2009,7(12):e1000253.
[7]Kuhné MR,Lin J,Yablonski D,et al.Linker for activation of T cells,zeta-associated protein-70,and Src homology 2 domain-containing leukocyte protein-76 are required for TCR-induced microtubule-organizing center polarization[J].J Immunol,2003,171(2):860-866.
[8]Glaser R,Andridge R,Yang EV,et al.Tumor site immune markers associated with risk for subsequent basal cell carcinomas[J].PLoS One,2011,6(9):e25160.
[9]Riccobon A,Gunelli R,Ridolfi R,et al.Immunosuppression in renal cancer:differential expression of signal transduction molecules in tumor-infiltrating,near-tumor tissue,and peripheral blood lymphocytes[J].Cancer Invest,2004,22(6):871-877.
[10]Hanaoka N,Jabri B,Dai Z,et al.NKG2D initiates caspase-mediated CD3ζ degradation and lymphocyte receptor impairments associated with human cancer and autoimmune disease[J].J Immunol,2010,185(10):5732-5742.
[11]Deng GM,Beltran J,Chen C,et al.T cell CD3ζ deficiency enables multiorgan tissue inflammation[J].J Immunol,2013,191(7):3563-3567.
[12]Nielsen SE,Zeuthen J,Lund B,et al.Phase I study of single,escalating doses of a superantigen-antibody fusion protein(PNU-214565)in patients with advanced colorectal or pancreatic carcinoma[J].J Immunother,2000,23 (1):146-153.
[13]Sundstedt A,Celander M,Hedlund G.Combining tumortargeted superantigens with interferon-alpha results in synergistic anti-tumor effects[J].Int Immunopharmacol,2008,8(3):442-452.
[14]高永鹏,林 晨,田红霞,等.BCR-ABL-SEA DNA疫苗诱导BALB/c小鼠的免疫应答[J].中国病理生理杂志,2011,27(2):361-366.
[15]徐水凌,毛亚飞,张梅光,等.葡萄球菌肠毒素A基因原核表达系统的构建及其表达产物的鉴定[J].中国病理生理杂志,2007,23(1):163-167.
[16]林 晨,高 珂,白 雪,等.超抗原SEA联合PMLRARα对外周血T细胞TCR Vβ亚家族基因表达的影响[J].免疫学杂志,2008,24(5):530-533.
[17]Gao H,Lee BN,Talpaz M,et al.Imatinib mesylate suppresses cytokine synthesis by activated CD4 T cells of patients with chronic myelogenous leukemia[J].Leukemia,2005,19(11):1905-1911.
[18]Pitcher LA,Mathis MA,Young JA,et al.The CD3 γε/δε signaling module provides normal T cell functions in the absence of the TCR ζ immunoreceptor tyrosine-based activation motifs[J].Eur J Immunol,2005,35(12):3643-3654.
Superantigen SEA antagonizes inhibitory effect of imatinib on T cell activation
YAN Yu-hui1,CHEN Xiao-hua1,WANG Guan-ming1,LIN Chen1,LI Yang-qiu2
(1Department of Microbiology and Immunology,School of Medicine,2Institute of Hematology,Jinan University,Guangzhou 510632,China.E-mail:tlinc@jnu.edu.cn)
AIM:To investigate the effects of staphylococcal enterotoxin A(SEA)on the inhibition of T lymphocyte activation induced by imatinib mesylate(IM).METHODS:Jurkat cells were stimulated with SEA(2 mg/L)and IM(5 nmol/L)for 24 h.The mRNA expression of CD3ε and ζ chains was measured by real-time fluorescence quantitative PCR.The protein levels of CD3ε and ζ chains were detected by Western blotting.RESULTS:The expression of CD3ε and ζ chains at mRNA and protein levels was down-regulated in the Jurkat cells stimulated by IM alone.These down-regulations of CD3ε and ζ chains were reversed by the stimulation of IM combined with SEA.The antagonistic effect of SEA on IM-mediated inhibition of CD3ε mRNA expression was significantly greater than that on CD3ζ mRNA.CONCLUSION:SEA antagonizes imatinib-mediated inhibitory effect on T cell activation.
Superantigen;Staphylococcal enterotoxin A;Imatinib;T-lymphocytes;Antigens,CD3
R364
A
10.3969/j.issn.1000-4718.2014.03.004
1000-4718(2014)03-0404-04
2013-10-21
2014-01-16
国家自然科学基金资助项目(No.81270604);广东省自然科学基金重点项目(No.S2013020012863);中央高校基本科研业务费专项资金资助项目(No.21612116)
△通讯作者Tel:020-85220257;E-mail:tlinc@jnu.edu.cn
猜你喜欢
猪业科学(2021年3期)2021-05-21
昆明医科大学学报(2021年1期)2021-02-07
中国医药指南(2019年14期)2019-01-07
农家科技下旬刊(2017年3期)2017-04-26
中国实用医药(2016年22期)2016-08-19
中国医药生物技术(2015年4期)2015-12-26
肿瘤影像学(2015年3期)2015-12-09
中国当代医药(2015年9期)2015-03-01
中国当代医药(2015年9期)2015-03-01
癌变·畸变·突变(2015年3期)2015-02-27
