
木聚糖酶及其在食品工业中的应用*
2013-11-21 10:02:00闵兆升郭会明顾承真洪厚胜
食品与发酵工业 2013年10期
闵兆升,郭会明,顾承真,洪厚胜,3
1(南京工业大学理学院,江苏南京,210009)2(南京工业大学生物与制药工程学院,江苏 南京,210009)3(南京汇科生物工程设备有限公司,江苏南京,210009)
半纤维素是植物组织中含量仅次于纤维素的一种极其丰富的可再生资源,约占植物干重的三分之一左右。与纤维素相比,半纤维素更容易被微生物降解利用。其中木聚糖为半纤维素的重要组成成分,是一种复杂的多聚五碳糖。木聚糖的分子结构变化范围比较大,这是由于木聚糖是由β-1,4-糖苷键与β-D-吡喃型木糖单元相连接构成主链,当其分支程度不同以及糖不同(戊糖和己糖)时,主链或侧链上就会带有多种不同的取代基,如图1所示[1]。因此,木聚糖的彻底降解需要多种酶的参与,其中最主要的就是我们通常所说的木聚糖酶(Xylanase,EC.3.2.1.8),其特指的是内切β-1,4-D-木聚糖酶。
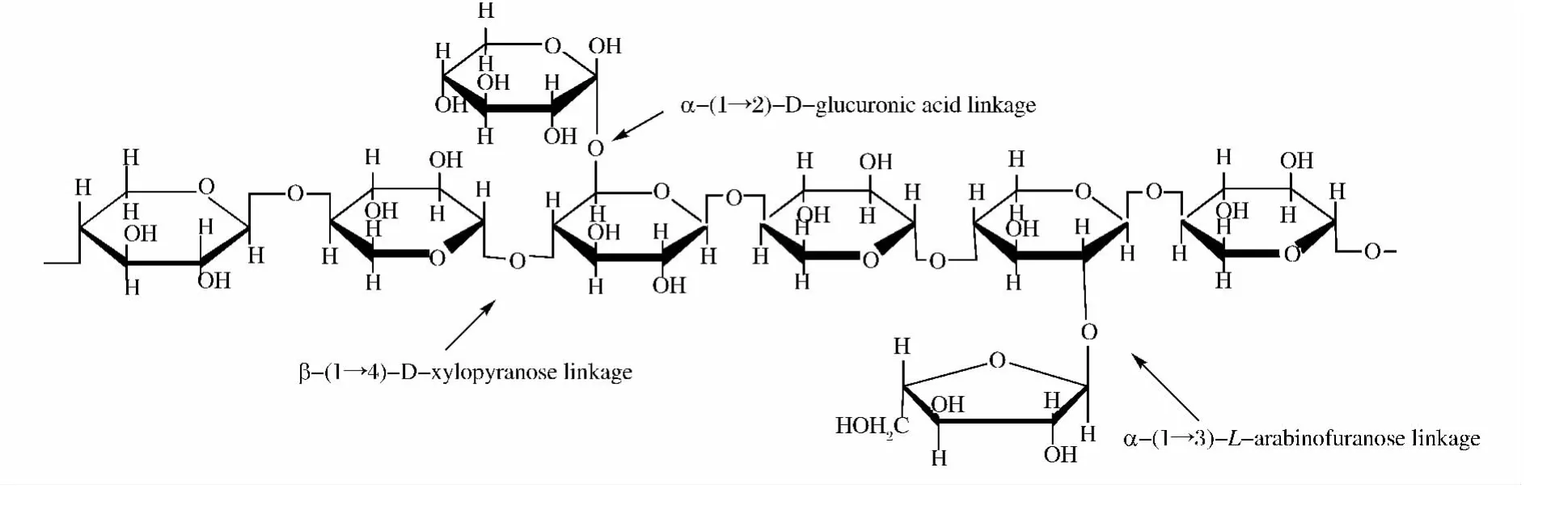
图1 连接不同侧链的木聚糖结构Fig.1 Xylan structure with different side chains attached
木聚糖酶(Xylanase,EC.3.2.1.8)广泛存在于自然界中,其主要来源为微生物,包括细菌、放线菌及真菌等。木聚糖酶通过随机的方式作用于木聚糖的主链,使其降解为低聚木糖和少量的木糖、阿拉伯糖。其在饲料以及造纸领域有着广泛的应用。在饲料业,木聚糖酶可作为饲用酶制剂,有效解除木聚糖的抗营养作用,促进畜禽对粗饲料的消化吸收;在造纸业,木聚糖酶可作为新型纸浆漂白助剂,解决传统的漂白工艺所带来的环境污染问题。另外,木聚糖酶还可以通过水解植物中的半纤维素,形成戊糖,进一步用于生产木糖醇、酒精、有机酸等产品,在可再生资源生物转化利用中也具有重要作用[2-3]。正是由于木聚糖酶所展现出的显著的研究价值和工业应用前景,使得越来越多的科研机构和人员投身于木聚糖酶的开发和研究工作。并且随着基因工程、蛋白质工程等高新生物技术的发展,关于木聚糖酶的基因克隆、表达和重组的报道也越来越多[4]。下面主要介绍一下木聚糖酶的特性、发酵生产及在食品工业中的应用。
1 木聚糖酶
广义的木聚糖酶是指能够降解半纤维素木聚糖的一组酶的总称,包括多种内切酶和外切酶。但通常所说的木聚糖酶仅限于β-1,4-D-木聚糖酶,其主要负责木聚糖主链骨架的降解,是木聚糖降解酶系中最关键的酶,也是当前木聚糖酶研究的热点。
1.1 木聚糖酶的结构与分类
木聚糖酶的结构比较复杂,有只含有单一区域即催化区的木聚糖酶,也有含有催化区和多种非催化区的木聚糖酶。其中,多区域木聚糖酶的分子结构中含有催化结构域(catalytic domain,CD)、纤维素结构域(cellulose binding domain,CBD)、木聚糖结构域(xylan-binding domain,XBD)、热稳定结构域、连接序列、重复序列和其他未知功能的非催化结构域。木聚糖酶催化区(CD)是酶催化水解的主要区域,并且可作为该酶的分类基础。不同来源的木聚糖酶,其氨基酸组成的数目和种类相差很大,但它们在催化区的大小都趋于一致。木聚糖酶的催化活性位点氨基酸与纤维素酶相似,主要为色氨酸和羧基氨基酸,有的还有半胱氨酸[5]。
以糖苷水解酶的催化结构域的氨基酸序列相似性和疏水簇分析法为基础,Hemnissat和Bairoch将糖基糖苷键水解酶中的内切木聚糖酶分为10(F)族和11(G)族,它们在催化特性、分子量、净电荷、酸碱性等方面均有比较大的差异。一般来说,F家族主要包括酸性的高分子量(>30 kDa)的内切木聚糖酶,而G家族则包含有略呈碱性的低分子量(<30 kDa)的内切木聚糖酶,且G家族的木聚糖酶对木聚糖具有很高的特异性。另外,属于同一家族的木聚糖酶催化区域具有同源性,因此可以根据同族的酶来推断未知酶的催化特性[6]。
1.2 木聚糖酶的理化性质
由于木聚糖酶结构的复杂性和来源的广泛性,使得微生物木聚糖酶具有多样性。同时,不同基因的产物,微生物所分泌蛋白酶的水解作用,以及糖基化和酰胺化等不同程度的化学修饰作用,都是造成微生物木聚糖酶多样性的原因。正是木聚糖酶的这种多样性,使得不同来源的木聚糖酶的组成和性质也不尽相同,因此关于木聚糖酶的理化性质研究更多的是集中于细菌和真菌产的木聚糖酶,主要性质有以下几点:①分子质量:多数木聚糖酶是单亚基蛋白,分子质量在8~45 kDa;②耐酸碱性:细菌所产木聚糖酶可大体分为2类:高分子量的耐酸木聚糖酶及低分子量的耐碱木聚糖酶。但在真菌中却没有这种差别,不过,低分子量木聚糖酶的耐碱性却是共同的。多数木聚糖酶的最适pH值为4~7,但是通常在pH值为8~10的范围内也能显示出很好的稳定性[6-7]。Taneja等[8]筛选到1株嗜碱真菌Aspergillus niduans.KK-99,该菌产的木聚糖酶的最适pH即为8.0。陈士成等[9]分离到1株短小芽孢杆菌Bacillus pumilus,其木聚糖酶的最适作用pH值为9.0。而杨革等[10]分离到1株细菌,其产生的木聚糖酶在pH 10.0左右才有最大酶活。木聚糖酶的这种宽范围的耐酸碱性,给木聚糖酶在更宽领域的工业应用提供了可能;③耐热性:多数细菌产的木聚糖酶的最适反应温度一般在40~60℃之间,真菌产的木聚糖酶的最适反应温度为50℃左右,反应温度都较为温和。一般来说,酶的热稳定性直接影响其应用领域,其耐热性直接影响其能否工业化生产。例如饲用木聚糖酶,既要求酶的最适温度与动物体内生理条件相近,又要有一定的热稳定性,以便于制粒干燥和包装储运;④等电点(pI):各种来源的木聚糖酶的等电点一般在3~10之间;⑤活性位点氨基酸组成:研究表明构成木聚糖酶活性区域的氨基酸主要包括天冬氨酸、谷氨酸、甘氨酸、丝氨酸和苏氨酸等。⑥糖类成分:来源于真核生物所产的木聚糖酶通常为糖基化酶,而来源于原核生物如链霉菌种、耐碱嗜温杆菌种等所产的木聚糖酶则为糖蛋白。这些糖类基因可与蛋白质共价连接或与木聚糖酶形成能够解离的复合体。研究表明,正是由于糖基化的作用才使得多糖酶能够在非常环境下也能保持较好的稳定性[11]。
1.3 木聚糖酶的诱导和调节机理
近年来,有研究表明,少数微生物木聚糖酶能够在没有外源诱导物的情况下组成性表达木聚糖酶,但是绝大多数的木聚糖酶还是诱导性表达酶。因此,除了菌种本身的特性以外,基质中的诱导物与酶的连接合成也有着密切的关联。通常来说,木聚糖酶的诱导过程是复杂的,同一诱导物,但是其作用的菌株不同,其产生的效果就可能非常大。对于某一种微生物来说,木糖可以诱导其高效表达木聚糖酶,产生最大的木聚糖酶活性,但对于另外一种微生物就有可能产木聚糖酶的抑制剂[12]。韩晓芳等[13]研究了 1株产木聚糖酶嗜碱芽抱杆菌,研究发现葡萄糖对产酶无抑制作用,并且木糖对该菌株产酶的诱导效果明显优于木聚糖。目前,关于木聚糖酶的诱导机制还不是很明确,较为普遍的一种观点是:低浓度的、小分子的低聚木糖等才能够对木聚糖酶的产生具有诱导作用。而木聚糖是一种大分子聚合物,不能直接穿过细胞膜进入细胞中,只有降解为小分子碎片后才能进入细胞内,从而进行木聚糖酶的诱导。图2显示了微生物细胞诱导产酶的机制。随着微生物细胞的生长,首先分泌出少量的组成型木聚糖酶,这些组成型木聚糖酶能够将木聚糖降解成木二糖、木糖和低聚木糖,而这些少量的降解产物能够先结合到细胞表面上,进而进入细胞内,启动转录加强酶的合成。因此,木聚糖酶诱导物的作用实际上就是它能够与对应的阻遏蛋白相结合,并且降低阻抑蛋白与操纵基因的结合力,从而使结构基因能转录生成对应的mRNA,进而产生对应的酶[12,14-15]。

图2 木聚糖酶诱导机制Fig.2 The mechanism of induction of xylanase
1.4 木聚糖酶的发酵生产
由于从自然界中分离得到的野生型产酶菌株,往往产酶能力低下,酶的纯度不高,稳定性也不够,在环境改变的情况下往往会大幅度丧失活性。所以工业上,木聚糖酶制剂的生产所用菌种几乎都是来自于通过基因构建开发出来的特殊工程菌。自20世纪80年代以来已有一百多种不同菌株的木聚糖酶基因被克隆并表达在合适的宿主中。其中研究和应用最多的当属毕赤酵母表达系统。这是因为酵母作为一种最简单的单细胞真核生物,兼有原核生物和真核生物的某些优点:酵母在简单培养基中便可快速生长,且能够进行细胞的高密度培养,易操作。另外,与大肠杆菌不同的是,毕赤酵母表达的外源蛋白常伴有糖基化现象,糖基化作用一方面能够通过影响蛋白质的构象起到增强木聚糖酶的稳定性作用,另一方面,能高效分泌表达外源蛋白,易于提纯,无内毒素污染,能够达到饲料、食品与医药行业应用的安全性要求。此外,酵母菌的规模化发酵工艺和下游处理工艺己相当成熟,利于木聚糖酶的大规模工业化生产[16-17]。目前,国内生产木聚糖酶的工艺便是基于重组毕赤酵母的高密度发酵技术。表1则列出了部分在毕赤酵母中表达的木聚糖酶基因。

表1 在毕赤酵母中表达的木聚糖酶基因Table 1 Expression of xylanase gene in Pichia pastoris
2 木聚糖酶在食品中的应用
2.1 木聚糖酶在面制品中的应用
目前,在食品行业中,应用木聚糖酶最多的当属在面包的烘焙过程中将其作为食品改良剂使用。制作面包的小麦面粉中含有少量以阿拉伯木聚糖(arabinoxylan,简称AX)为主的戊聚糖,其含量约为2%~3%[24]。根据阿拉伯木聚糖在水中的溶解性可将其分为:水溶性阿拉伯木聚糖(water-extractablearabinoxylan,简称WEAX)和水不溶性阿拉伯木聚糖(water-unextractable anrabinoxylan,简称 WUAX),其中WEAX约占25% ~30%[25]。虽然小麦面粉中AX的含量并不高,但其吸水性强,对面团的形成及面包品质有重要影响。张勤良等[26]的研究证明,在面包的制作过程中,适当添加一定量的木聚糖酶,能够改善面团的弹性、稳定性和对过度发酵的耐受性,使得面粉的粉质,面团的揉制都得到了改善。李秀婷等[27]的研究也证明了,添加适当剂量的木聚糖酶后的面包,在烘焙过后不仅质地洁白、组织细腻、气孔均匀、入口松软有嚼劲且过早老化的问题也得到了改善。
2.1.1 木聚糖酶对面包制作过程中面团品质的影响
在面团揉制过程中,木聚糖酶能够促使部分的WUAX溶解,其使得面团的持水能力下降,从而促使面团中其他组分中的水分重新得到分布,进而可增加面团的延伸性。而面团延伸性的增强也会反过来增加面团水相的黏度,面团的黏度上升又会使得面团的内聚力增强,弹性增大。但是,过量的木聚糖酶会使面团中的水分过多释放而影响面团的质量。这是因为在面包的揉制过程中,WUAX的增溶速率最快,WEAX过度降解,进而使得面团的内聚力下降,黏度过高,稠度下降,同时面团的抗延伸性也下降,进而影响了面包的揉制和制作。因此,在面包的制作过程中,要控制好木聚糖酶的添加量。另外,面团醒发对于面包制作具有重要的影响,但是目前关于这方面的研究和报道都很少,主要是因为面团醒发过程中面团的产气能力、持气能力以及面团的稳定性很难测定[26,28]。
2.1.2 木聚糖酶对面包在烘焙过程中的影响
面包烘焙的开始阶段可以被认为是面团醒发的外延,此时,由于气体的膨胀和酵母活性的高效发挥,使得面团膨胀很快。因此,这一阶段,面筋的黏度、弹性以及面团的持气能力都要比醒发过程显得更为重要。面粉中的WEAX在烘焙初期可以增加气泡的强度和稳定性,改善面包心的均匀性,对面包的烘焙有着非常积极的作用;但是,WUAX对面包的烘焙初期却具有负面影响,它会降低面包的体积,破坏面包心的均匀性和面包的质构。然而,在烘焙过程中,添加适当剂量的木聚糖酶就能够使得面包体积增加、质构改善、面包心的均匀性提高[26,29]。其原因包括以下几点:第一,木聚糖酶能够促使WUAX增溶;第二,WEAX的适当降解,使附着在面筋蛋白质膜表面的AX黏度下降,从而促使面筋蛋白质膜延伸性增加,但是面团的稳定性和面筋的网络结构并未因此受到破坏;第三,不断增加的WEAX与面包中的氧化剂发生了氧化凝胶作用,使烘焙的面包持水性加强,弹性增加。
2.1.3 木聚糖酶对面包抗老化的影响
面包老化是指面包皮的破坏、风味的降低、面包变硬等现象,其主要原因是面包在贮藏过程中淀粉发生了凝沉。但是当添加适当剂量的木聚糖酶后,能够使面包中的WUAX和WEAX的比例更加合理,而且WEAX的分子链长度也有所下降。因为WEAX能够与面包其他组分中的氧化剂发生氧化胶凝作用,增加了面包的持水性;而WEAX的链长下降可以降低WEAX与邻近生物大分子之间链的连接,使起水桥作用的水分子的量减少,这样就能有效延缓面包的失水速率和水分迁移速率,进而使得面包抗老化能力增强[27,30]。
2.2 木聚糖酶在低聚木糖制备中的应用
低聚木糖是一种功能性低聚糖。低聚木糖对pH值及热的稳定性较好,因此其在人体内难以消化,但是却可以极好地促进肠道内双歧杆菌的增殖,从而起到抑制肠道内腐败菌的生长,改善肠道微生态环境,调节人体的生理机能,增强免疫力,预防疾病发生的作用;另外,由于低聚木糖的甜度仅为蔗糖的40%,因此,食用该糖后不会导致人体血液中葡萄糖水平大幅度上升,从而可以作为功能性甜味剂,部分取代蔗糖,给糖尿病患者食用;再者,低聚木糖具有良好的持水性,在食品中可作为保湿剂或保鲜剂使用[31-32]。
目前,工业制备低聚木糖最常用的方法就是酶解法,即使用微生物产生的内切型木聚糖酶分解木聚糖,然后经分离提纯制得低聚木糖,由于是利用内切型木聚糖酶定向酶解半纤维素,故副产物较少,有利于后续工艺中低聚木糖的分离、提纯和精制。在我国,杨瑞金等[32]最早研究并报道了酶法生产低聚木糖,他们是利用P-3-31菌株的木聚糖酶酶系将玉米芯中的木聚糖水解制取低聚木糖。制备低聚木糖的木聚糖酶是有特殊要求的:首先,对底物水解的特异性要求:即要求产生木二糖、木三糖的比例要高;其次需要不含有木糖苷酶活性。目前,关于木聚糖酶水解底物的特异性及其作用模式在国外仍是研究热点,这也是制备低聚木糖的关键技术。
2.3 木聚糖酶在酿酒过程中的应用
目前,用于酿酒行业的木聚糖酶为酸性木聚糖酶,原因在于木聚糖酶对谷物细胞壁中木聚糖的作用有助于加快水解淀粉酶,其主要应用对象也是指啤酒和清酒的酿制。但基于成本的考虑,至今为止,在酿酒行业大量使用木聚糖酶还不常见,关于这方面的研究也鲜有报道。但是早在1991年,日本就已经有了将木聚糖酶用于日本烧酒生产中的应用研究,并且取得了比较好的结果[33]。我国江南大学陆健等[34]也对耐酸性木聚糖酶应用于清酒的酿造作了研究,结果发现,添加木聚糖酶,既提高了发酵速度,又提高了酒精产率。但是要想在酿酒工业实现实际的工业化生产,就必须满足以下条件:首先要求使用的木聚糖酶能够耐酸性,以适应实际的生产条件;其次该酶应该有比较好的底物水解针对性,能够快而有效地分解谷物细胞壁中的木聚糖,从而加快淀粉酶的作用;再者,木聚糖酶的产率要高,这样才能够有经济上的可行性来将其应用于酿酒工业中。
3 结束语
由此可见,木聚糖酶在食品行业中有着广泛的应用前景,但是与其良好的水解能力和巨大的应用价值相比较而言,木聚糖酶的工业化生产及应用现状却依旧问题重重。首先是酶的稳定性问题。木聚糖酶是一种微生物制剂,对外界物化条件的影响极为敏感。然而应用在食品领域中的酶,往往需要经过高温的加工处理,这常会使酶遭到破坏,影响木聚糖酶活性的发挥,在一定程度上限制了木聚糖酶在食品领域的应用。其次是酶的添加量问题。不同性质和酶活的产品,都有其适合的应用领域,且在不同的应用条件下,即使在同一应用领域中,木聚糖酶的添加量和添加水平也存在一定的差异。因此,在实际应用中,如果使用不当,不但会影响酶的应用效果,还会适得其反。再者便是与其他酶制剂的复配问题。在实际应用中,往往需要添加其他酶系如果胶酶、纤维素酶等组成复合酶制剂用于生产,然而,不同组合及配比的复合酶制剂的应用效果是有差异的。因此,要解决好上述问题,就需要更加系统、深入地开展木聚糖酶应用研究,根据特定的应用对象有针对性地创造最佳的反应条件,最大限度地发挥木聚糖酶的作用,提高产品质量,降低生产成本,实现可持续发展。
[1]朱静,严自正.微生物产生的木聚糖酶的功能和应用[J].生物工程学报,1996,12(4):375-378.
[2]石军,陈安国.木聚糖酶的应用研究进展[J].中国饲料,2002(4):10-12.
[3]曲音波,高培基,陈嘉川.制浆造纸用木聚糖酶的研究进展[J].生物工程进展,1998,18(6):35-39.
[4]张红莲,姚斌,范云六.木聚糖酶的分子生物学及其应用[J].生物技术通报,2002(3):23-26+30.
[5]刘瑞田,曲音波.木聚糖酶分子的结构区域[J].生物工程进展,1998,18(6):25-27.
[6]孙振涛,赵祥颖,刘建军,等.微生物木聚糖酶及其应用[J].生物技术,2007,17(2):93-97.
[7]陆健,曹钰,陈坚,等.木聚糖酶的产生、性质和应用[J].酿酒,2001,28(6):30-34.
[8]Taneja K,Gupta S,Chander Kuhad R.Properties and application of a partially purified alkaline xylanase from an alkalophilic fungus Aspergillus niduans KK-99[J].Bioresource Technology,2002,85(1):39-42.
[9]陈士成,曲音波,刘相梅,等.短小芽孢杆菌A-30耐碱性木聚糖酶的纯化及性质研究[J].中国生物化学与分子生物学报,2000,16(5):698-701.
[10]杨革,李亚.细菌耐碱性木聚糖酶产生菌的选育及其酶学性质的初步研究[J].南京农业大学学报,1999,22(1):114-115.
[11]陆健,曹钰,陈坚,等.木聚糖酶的产生、性质和应用[J].酿酒,2001,28(6):30-34.
[12]江正强.微生物木聚糖酶的生产及其在食品工业中应用的研究进展[J].中国食品学报,2005,5(1):4-12.
[13]韩晓芳,郑连爽.产木聚糖酶嗜碱细菌的筛选及产酶条件研究[J].环境污染治理技术与设备,2002,3(11):25-27.
[14]岳晓禹,贺小营,牛天贵,等.木聚糖酶的研究进展[J].酿酒科技,2007(4):113-115,120.
[15]刘超纲,勇强,余世袁.纤维素和木聚糖复合诱导合成木聚糖酶的研究[J].林产化学与工业,2001,21(1):69-73.
[16]Thongekkaew J,Ikeda H,Masaki K,et al.An acidic and thermostable carboxymethyl cellulase from the yeast Cryptococcus sp.S-2:Purification,characterization and improvement of its recombinant enzyme production by high cell-density fermentation of Pichia pastoris[J].Protein Expression and Purification,2008,60(2):140-146.
[17]张红莲.橄榄绿链霉菌Streptomyces olivaceoviridis A1所产木聚糖酶的纯化、酶基因的克隆及表达[D].北京:中国农业科学院饲料研究所,2002.
[18]苏玉春.木聚糖酶的酶学特性及基因克隆表达研究[D].长春:吉林农业大学,2008.
[19]Chen X,Xu S,Zhu M.Site-directed mutagenesis of an Aspergillus niger xylanase B andits expression,purification and enzymatic characterization in Pichia pastoris[J].Process Biochemistry,2010,45(1):75-80.
[20]Jeya M,Thiagarajan S,Lee J.Cloning and expression of GH11 xylanase gene from Aspergillus fumigatus MKU1 in Pichia pastoris[J].Journal of Bioscience and Bioengineering,2009,108(1):24-29.
[21]何军.高效木聚糖酶基因工程菌的构建及重组酶酶学性质与功效研究[D].雅安:四川农业大学,2008.
[22]Ruanglek V,Sriprang R.Cloning,expression,characterization,and high cell-densityproduction of recombinant endo-1,4-β-xylanase from Aspergillus niger in Pichia pastoris[J].Enzyme and Microbial Technology,2007,41(1-2):19-25.
[23]Chantasingh D,Pootanakit K,Champreda V.Cloning,expression,and characterizationof a xylanase 10 from Aspergillus terreus(BCC129)in Pichia pastoris[J].Protein Expressionand Puri Wcation,2006,46(1):143-149.
[24]Courtin C M,Delcour J A.Arabinoxylans and endoxylanases in wheat flour bread-making[J].Journal of Cereal Science,2002,35(3):225-243.
[25]周素梅,向波,王璋,等.小麦面粉中阿拉伯木聚糖研究进展[J].粮油食品科技,2001,9(2):20-22.
[26]张勤良,王璋,许时婴.中性木聚糖酶在面包制作中的应用[J].食品与发酵工业,2004,30(7):21-25.
[27]李秀婷,李里特,江正强,等.耐热木聚糖酶对面包老化作用探讨[J].食品与发酵工业,2006,32(1):23-27.
[28]Caballero P A,Gómez M,Rosell C M.Improvement of dough rheology,bread quality and bread shelf-life by enzymes combination[J].Journal of Food Engineering,2007,81(1):42-53.
[29]Ahmad Z,Butt M S,Ahmed A,et al.Effect of Aspergillus niger xylanase on dough characteristics and bread quality attributes[J].Journal of Food Science and Technology,2012:1-9.
[30]Driss D,Bhiri F,Siela M,et al.Improvement of breadmaking quality by xylanase GH11 from Penicillium occitanis Pol6[J].Journal of Texture Studies,2012,44(1):75-84.
[31]郑建仙,耿立萍.功能性低聚糖析论[J].食品与发酵工业,1997,23(1):39-46.
[32]杨瑞金,许时婴,王璋.低聚木糖的功能性质与酶法生产[J].中国食品添加剂,2000,(02):89-93.
[33]Ogasawara H.Contribution of hemicellulase in Shochu koji to the resolution of barley in Shochu mash[J].J Brew Soc Jpn,1991,86:304-307.
[34]陆健,曹钰,陈坚,等.耐酸性木聚糖酶在清酒酿造中的作用[J].食品与发酵工业,2002,28(1):27-30.
猜你喜欢
阅读(中年级)(2022年9期)2022-10-08 01:56:14
林产化学与工业(2022年4期)2022-09-13 09:47:12
红蜻蜓·低年级(2022年2期)2022-03-19 22:44:41
娃娃乐园·综合智能(2022年1期)2022-01-19 10:12:26
红蜻蜓·低年级(2021年12期)2021-12-19 15:06:23
湖南饲料(2019年5期)2019-10-15 08:59:10
学苑创造·A版(2017年12期)2018-01-17 19:05:54
小学生导刊(2017年34期)2017-07-09 03:00:58
中国酿造(2016年12期)2016-03-01 03:08:21
广东饲料(2016年8期)2016-02-27 11:10:01
