
太原汉族人群17个Y-STR基因座等位基因频率及遗传关系
2013-11-15 02:56陈惠珍张娟层刘金杰薛淑莲郭大玮梁景青太原市公安局刑侦支队技术处DNA室太原0000山西医科大学法医学院山西医科大学公共卫生学院流行病教研室通讯作者mailliangjingqingsinacom
山西医科大学学报 2013年10期
陈惠珍,张 戎,张娟层,刘金杰,薛淑莲,郭大玮,梁景青(太原市公安局刑侦支队技术处DNA室,太原 0000;山西医科大学法医学院;山西医科大学公共卫生学院流行病教研室;通讯作者,E-mail:liangjingqing@sina.com)
人类Y染色体属于性染色体,除拟常染色区外,在遗传过程中不发生交换重组,其序列结构特征能稳定地由父亲传给儿子,呈父系遗传。因此,YSTR基因座在法医学个体识别、亲子鉴定、混合斑中男性成分的检测、追溯父系迁移历史等方面都具有独特的应用价值。本课题旨在为太原汉族人群YSTR单倍型提供基本数据资料,通过应用Y-filerTM试剂盒,对17个Y-STR基因座进行分析和研究,结合其他地区人群样本资料,分析遗传距离和遗传关系,为太原汉族人群的历史迁移提供生物学的支持。
1 材料和方法
1.1 样本采集
选取居住在太原地区的102例汉族无关男性个体的血样,样本系太原市公安局DNA实验室日常案件积累。
1.2 Y-STR 基因座的选取
本课题应用美国ABI公司的Y-filer试剂盒中17个Y-STR基因座进行研究。
1.3 5%Chelex-100快速提取法提取DNA
1.3.1 检材预处理 剪取1 cm×1 cm大小的血卡,置于1.5 ml的 EP管中,加入1 000 μl超纯水,震荡混匀后14 000 r/min离心3 min,弃上清,收集沉淀。1.3.2 提取 DNA 每个 EP管中加入200 μl 5%Chelex-100和20 μl蛋白酶K,震荡混匀后,在56℃水浴箱中孵育40 min。置于100℃加热模块中,变性8 min,14 000 r/min 离心3 min,取出,静置。
1.4 PCR的扩增及扩增产物的检测
1.4.1 PCR扩增 PCR扩增在ABI 9700扩增仪上进行,10 μl扩增体系包括:Amp FlSTR® Y-filer Kit Primer Set 2 μl,Y-filer PCR Reaction mix 2.5 μl,Ampli Taq GoldTMDNA(5 U/L)0.2 μl,DNA 模板 0.8 μl,加去离子水至10 μl。PCR热循环参数:95℃,11 min;94℃,1 min;61℃,1 min;72℃,1 min;30个循环;60℃,80 min;4℃保存。
1.4.2 扩增片段的电泳分离与检测 采用 ABI 3130xl型基因分析仪进行毛细管电泳分离,应用GeneMapper IDv 3.1软件进行基因分型。
1.5 数据分析
各基因座等位基因频率采用直接计数法,各基因座基因变异度(GD),根据公式GD=[n(1-∑Pi2)]/(n-1)计算,其中Pi为等位基因的频率,n为样本数[1,2];计算累积的 GD 值(TGD),按公式TGD=1-(1-GD1)(1-GD2)(1-GD3)(1-GD4)…(1-GDn)计算,GDn为各基因座的GD值。登陆单倍型数据库(www.yhrd.org),计算群体间的遗传距离Rst矩阵。用MEGA5.05软件建立群体相邻连接(neighbor-joining,NJ)系统发生树。
102 例太原汉族人群数据与世界各群体共享信息:登陆单倍型数据库(www.yhrd.org),按照DYS19,DYS389Ⅰ,DYS389Ⅱ,DYS390,DYS391,DYS392,DYS393,DYS385a,DYS385b,DYS438,DYS439,DYS437,DYS448,DYS456,DYS458,DYS635,Y-GATA-H4的顺序,依次输入102例太原汉族人群数据资料,计算太原汉族人群与世界各个群体间的相同概率。
2 结果
2.1 102例太原汉族无关男性群体17个Y-STR基因座遗传多态性
在102例太原汉族无关男性群体个体中,17个Y-STR基因座均得到有效的扩增。DYS385a/b基因座共检出41种单倍型,其余15个基因座共检出84个等位基因,等位基因频率在0.009 8-0.725 5之间,基因多样性在0.433 5-0.894 0之间,除了DYS391、DYS437、DYS438,其余基因座的基因多样性均大于0.5(见表1)。该结果说明:17个Y-STR基因座在山西太原汉族人群中有较好的遗传多态性,且有较强的个体识别能力及非父排除能力,适合法医学的实际应用。
2.2 太原汉族人群与9个少数民族群体间的遗传距离
本实验研究发现,太原地区汉族人群基因座等位基因及频率分布与国内其他地区的少数民族人群相比,具有较大的差异性(见表2)。太原汉族与其他9个[3-11]少数民族群体的遗传距离都较大,其中,与辽宁满族人群之间的遗传距离最小(0.001 4),与台湾地区泰雅族之间的遗传距离最大(0.469 6)。
2.3 太原汉族人群与10个群体间的遗传距离
本实验研究发现,太原地区汉族人群中少数基因座等位基因及频率分布与国内其他地区的人群相比[12-15],具有一定的差异性(见表3)。数据结果表明:太原汉族与北京汉族之间的遗传距离最小(0.001 7),与台湾泰雅族人群之间的距离最大(0.469 6)。
2.4 太原汉族人群与11个群体间的N-J系统发生树
2.4.1 太原汉族人群与9个少数民族群体间的NJ系统发生树 太原汉族与9个少数民族间的N-J系统发生树结果显示:汉族人群与少数民族人群之间有一定的遗传距离。太原汉族和辽宁满族、北京汉族聚为一类,与广西壮族和福州畲族遗传距离较近;卫藏地区藏族、安多地区藏族、康巴地区藏族和青海藏族聚为一大类,台湾泰雅族和台湾排湾族聚为一类(见图1)。
2.4.2 太原汉族人群与10个群体间的N-J系统发生树 太原汉族与10个群体间的N-J系统发生树结果显示:太原汉族与西安汉族遗传距离最近,与北京汉族聚为一类;河南汉族与广西壮族、云南汉族聚为一大类;安多地区藏族和卫藏地区藏族聚为一类;浙江汉族,代表南方汉族人群单独为一类;台湾泰雅族和台湾排湾族聚为一类(见图2)。
3 讨论
3.1 太原汉族人群17个Y-STR基因座等位基因频率及遗传多态性

表1 太原市汉族无关男性个体17个Y-STR基因座等位基因频率分布及GD值 (n=102)Table 1 Allelic frequencies and gene diversity of 17 Y-STR loci in Taiyuan Han populations (n=102)

表2 个不同种族群体间Rst值的遗传距离矩阵Table 2 The genetic distance between 11 populations
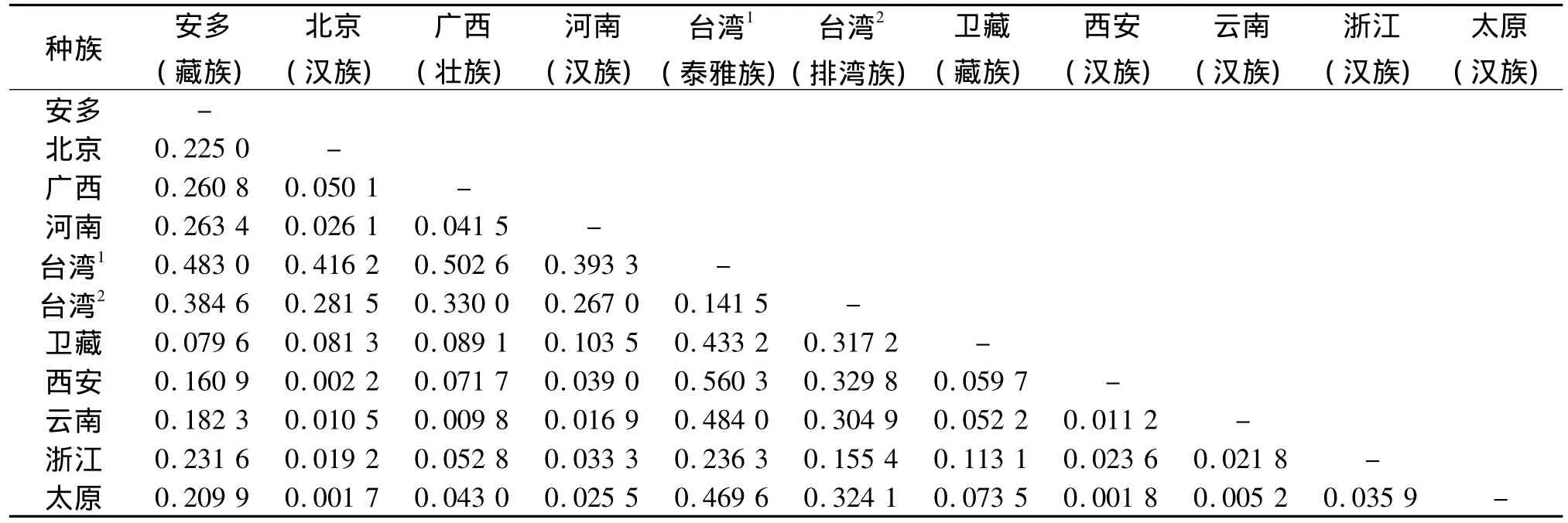
表3 11个不同群体间Rst值的遗传距离矩阵Table 3 The genetic distance between 11 populations

图1 太原汉族与9个少数民族间的N-J系统发生树Figure 1 Neighbor-joining tree between Taiyuan Han population and nine minority groups

图2 太原汉族与10个群体间的N-J系统发生树Figure 2 Neighbor-joining tree between Taiyuan Han population and ten populations
对102例太原汉族无关男性群体个体的研究发现,17个 Y-STR 基因座 DYS456、DYS390、DYS389Ⅰ、DYS389Ⅱ、DYS458、DYS19、DYS393、DYS391、DYS439、 DYS635、 DYS392、 DYS437、 DYS438、DYS448、Y-GATA-H4 基因座分别检出 5、6、4、7、7、5、5、5、6、6、7、4、6、7、4 个等位基因;DYS385a/b 基因座等位基因频率在0.009 8-0.725 5之间,基因多样性在 0.433 5-0.894 0之间,除了 DYS391、DYS437、DYS438,其余基因座的基因多样性均大于0.5。说明17个Y-STR基因座在山西太原汉族人群中有很好的遗传多态性,适合法医学的实际应用。
根据N-J系统发生树的结果,我们可以发现:汉族人群与少数民族群体间有一定的遗传距离,而少数民族中的满族人群和壮族人群与汉族人群遗传距离较近,这与我国的历史条件和地理环境有密切关系。以浙江为代表的南方汉族人群与北方汉族人群间有较远的遗传距离;藏族人群和台湾人群各自聚为一类。
3.2 太原地区汉族人群和其他群体遗传关系的分析
Y-STR基因座有父系遗传特征,只能由父亲传递给儿子[16],且在传递过程中是以较稳定的单倍体形式。由于地理、婚配以及文化等方面的原因,YSTR单倍型的基因频率有较明显的资料差异。山西太原是山西省的省会,在2 500年的历史长河中,太原曾经是唐尧故地、战国名城、太原故国、北朝霸府、天王北都、中原北门、九边重镇、晋商故里,“无端更渡桑干水,却望并州是故乡”(摘自贾岛的《渡桑干》。明初,大量山西人群持续50余年大规模强制性移民至北京、河北、陕西、河南等华北平原地区,给他们的后裔留下了许多独特的生活习俗和生理特征。除此之外,在山西历史上有浓重一笔的俗称“走西口”和“闯关东”,也使山西向东北、内蒙古和其他地区输出一部分移民。与此同时,山西地区也接收了大量的来自全国的外来移民,省会太原更是外来移民首选的移民地,经过多年的发展,与本地人融合,成为一体,组成了现代的太原人群。
通过本实验,发现太原汉族人群与北京汉族之间的遗传距离最小(0.001 7),其次是与西安汉族之间的遗传距离(0.001 8)。这些数据都说明历史上山西人群的大规模迁移,与这些地方的人群的融合有很大的影响。
本研究所研究的17个Y-STR基因座组成的单倍型,其计算的遗传距离结果和山西汉族群体形成的历史过程基本一致,从基因座方向为山西太原的历史研究提供了有力的生物学证据。
[1] Gill P,Brenner C,Brinkmann B,et al.DNA Commissions of the ISFG:recommendations on forensic analysis using Y-Chromosome STRs[J].Int J Legal Med,2001,114:305-309.
[2] Gusmao L,Butler JM,Carraeedo A,et al.DNA Commission of the International Society of Forensic Genetics(ISFG):an update of the recommendations on the use of Y-STRs in forensic analysis[J].Int J Legal Med,2006,120(4):191-200.
[3] Gayden T,Bukhari A,Chennakrishnaiah S,et al.Y-chromosomal microsatellite diversity in three culturally defined regions of historical Tibet[J].Forensic Sci Int Genet,2012,6(4):437-446.
[4] 李斌,吕政,王澍,等.福建汉族人群17个Y-STR基因座遗传多态性调查[J].刑事技术,2008(2):60-62.
[5] 鲁涤,袁丽,杨雪.北京汉族群体17个Y-STR基因座遗传多态性[J].中国法医学杂志,2010,25(6):432-433.
[6] 滕少康,曹林枝,黄世宁,等.广西黑衣壮族17个Y-STR位点遗传多态性[J].基础医学与临床,2009,29(7):742-748.
[7] 冯东亮,刘长晖,梁祚仁,等.广西4个少数民族17个Y-STR基因座的多态性分析[J].遗传,2009,31(9):921-935.
[8] Zhu B,Wu Y,Shen C,et al.Genetic analysis of 17 Y-chromosomal STRs haplotypes of Chinese Tibetan ethnic group residing in Qinghai province of China[J].Forensic Sci Int,2008,175(2-3):238-243.
[9] Wu FC,Ho CW,Pu CE,et al.Genetic polymorphisms of 17 Y-chromosomal short tandem repeat loci in Atayal population of Taiwan[J].Croat Med J,2009,50(3):313-320.
[10] Wu FC,Ho CW,Pu CE,et al.Y-chromosomal STRs haplotypes in the Taiwanese Paiwan population[J].Int J Legal Med,2011,125(1):39-43.
[11] You XQ,Fen J,Zou QL.Y chromosome relative length analysis of 106 case of Fujian she ethnic[J].Yi Chuan,2002,24(3):257-258.
[12] Wu W,Pan L,Hao H,et al.Population genetics of 17 Y-STR loci in a large Chinese Han population from Zhejiang Province,Eastern China[J].Forensic Sci Int Genet,2011,5(1):11-13.
[13] Yanmei Y,Tao G,Yubao Z,et al.Genetic polymorphism of 11 Y-chromosomal STR loci in Yunnan Han Chinese[J].Forensic Sci Int Genet,2009,4(2):67-69.
[14] 郭利红,田曾元,刘亚举,等.河南汉族人群16个Y-STR基因座遗传多态性[J].中国法医学杂志,2010,25(2):66-67.
[15] 杨立,蔡继峰,常云峰,等.湖南汉族人群16个Y-STR基因座遗传多态性[J].中国法医学杂志,2011,26(2):136-137.
[16] 侯一平.法医物证学[M].2版.北京:人民卫生出版社,2005:95-96.
猜你喜欢
支部建设(2021年18期)2021-08-20
发明与创新(2021年17期)2021-07-05
医学与法学(2020年3期)2020-09-18
意林(儿童绘本)(2020年1期)2020-02-14
新生代·下半月(2019年7期)2019-09-10
中国现当代社会文化访谈录(2016年0期)2016-09-26
食管疾病(2015年3期)2015-12-05
地方文化研究(2015年4期)2015-11-07
新疆大学学报(哲学社会科学版)(2015年4期)2015-10-12
现代企业(2015年5期)2015-02-28
