
人EYA3基因真核表达载体的构建及其活性检测
2013-10-29 09:36符静徐小洁刘家宏田沛荣吕朝晖王涛朱建华陆菊明叶棋浓
生物技术通讯 2013年5期
符静 ,徐小洁 ,刘家宏 ,田沛荣 ,吕朝晖 ,王涛 ,朱建华 ,陆菊明 ,叶棋浓
1.军事医学科学院 生物工程研究所,北京 100850;2.解放军总医院 内分泌科,北京 100853
EYA(eyes absents)是一个进化上较保守的基因家族,属于视网膜决定基因网络成员[1],目前发现人EYA基因家族有4个成员即EYA1~EYA4。EYA基因表达的蛋白是一类具有酪氨酸磷酸酶活性的转录调控因子[2],这些因子不仅在胚胎发育时组织器官的特异性分化中起关键调控作用,而且还参与多种恶性肿瘤的发生发展。为了深入研究EYA3在乳腺癌中的作用,我们从人乳腺文库中扩增出EYA3基因的全长编码序列,构建在带myc标签的真核表达载体上,通过生长曲线实验验证该重组质粒能促进乳腺癌细胞的生长,证明其蛋白结构正确且具有活性,为今后的研究奠定了基础。
1 材料和方法
1.1 材料
人ZR75-1细胞由本实验室传代培养;乳腺文库和pXJ-40-myc载体为本室保存;VigoFect为威格拉斯生物技术有限公司产品;限制性内切酶、DNA连接酶、PCR试剂均购自TaKaRa公司;质粒提取、胶回收、PCR回收试剂盒购自Promega公司;DMEM及小牛血清均购自Gibco公司;测序由北京奥科生物技术有限责任公司完成。
1.2 myc-EYA3重组质粒的构建与测序
根据文献报道的人EYA3基因序列设计上游引物(5'-CCGCTCGAGATGGAAGAAGAGCAAGATTT AC-3')和下游引物(5'-GGGGTACCTTAGAGAAAA TCAAGCTCTAAAG-3'),其中上游引物含XhoⅠ酶切位点,下游引物含KpnⅠ酶切位点(下划线序列)。以人乳腺文库为模板,PCR扩增目的片段(95℃预变性5 min;95℃变性30 s,60℃退火30 s,72℃延伸90 s,31个循环;72℃延长7 min),用胶回收试剂盒回收PCR产物。
用XhoⅠ和KpnⅠ双酶切myc载体,经10 g/L琼脂糖凝胶电泳后,胶回收载体大片段;将PCR片段回收后再用XhoⅠ和KpnⅠ酶切,形成带有粘端的双链,用T4DNA连接酶连接入myc载体中;转化大肠杆菌DH5α,挑选克隆,振荡培养并提质粒,用XhoⅠ和KpnⅠ双酶切鉴定,酶切鉴定正确的克隆送北京奥科生物公司测序。
1.3 哺乳动物细胞转染
用不含双抗、含10%胎牛血清的DMEM培养基将ZR75-1细胞接种于6 cm皿中,接种量以转染时细胞密度达80%为宜,培养24 h后进行转染,转染前 1 h 换液。将 4 μL VigoFect与 200 μL NaCl混合,再将总量为 10 μg的重组质粒与 200 μL NaCl混合,然后将上述2种溶液轻轻混合,室温放置15 min,加入6 cm皿中,并以同样方法转染空myc载体作为对照,37℃、50%CO2常规培养,4~6 h换液。
1.4 Western印迹
质粒转染ZR75-1细胞24 h后收集细胞蛋白,加入2×SDS加样缓冲液,煮沸10 min,高速离心2 min,取上清液进行SDS-PAGE后电转移至硝酸纤维素膜上;用5%脱脂奶粉于4℃封闭过夜,加入用5%脱脂奶粉以1∶5000稀释的用HRP标记的抗myc标签鼠单克隆抗体,室温轻摇1 h,TBST洗膜3次,每次5 min,用化学发光法显色5 min,压片显影。
1.5 生长曲线实验
将myc-EYA3和空载体分别转染ZR75-1细胞,48 h后取处于对数生长期的细胞培植成单细胞悬液,细胞计数后稀释至2×104/L,分别接种于5块96孔板,各设3个重复孔,常规培养。每日取一块96孔板,在接种孔内用排枪加入10 μL CCK-8溶液,37℃放置1~4 h,测定D450nm值。
2 结果
2.1 myc-EYA3重组质粒的构建与鉴定
以实验室保存的乳腺文库为模板,PCR扩增人EYA3的编码序列,获得约1700 bp的DNA片段,与预期一致(图1)。将PCR产物用XhoⅠ和KpnⅠ双酶切后与同样经这2种酶酶切的myc载体连接,转化大肠杆菌DH5α,挑选克隆进行菌液PCR鉴定,若获得与目的条带1700 bp接近(图2)的克隆,则初步认为是带有人EYA3基因的阳性重组克隆。将所得阳性克隆提质粒经酶切鉴定,可切出2条长度分别约为5000和1700 bp的条带,而相应的空载体酶切只见大片段,符合预期结果(图3)。DNA序列测序结果表明,插入片段的DNA序列与人EYA3基因的编码序列完全一致(数据略)。
2.2 Western印迹检测myc-EYA3在ZR75-1细胞中的表达
将构建的myc-EYA3重组质粒转染ZR75-1细胞系,24 h后提取蛋白进行SDS-PAGE,Western印迹检测myc-EYA3蛋白的表达。结果见图4,用myc-HRP抗体能在相对分子质量约60×103处检测到明显特异性条带,说明myc-EYA3重组蛋白在ZR75-1细胞中能够正确表达,而空载体无表达。

图1 PCR扩增人EYA3的编码序列

图2 重组质粒myc-EYA3的菌液PCR电泳图谱

图3 重组质粒myc-EYA3的XhoⅠ/KpnⅠ双酶切电泳图谱
2.3 生长曲线实验验证EYA3促进ZR75-1细胞生长的功能
为验证EYA3的功能,我们将真核表达载体myc-EYA3及空载体转染ZR75-1细胞后进行生长曲线实验,结果证明EYA3能促进ZR75-1细胞的生长(图5)。
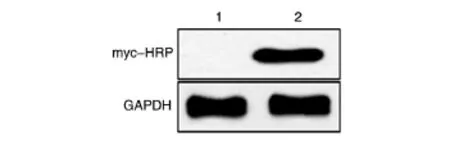
图4 Western印迹检测融合蛋白的表达

图5 EYA3促进ZR75-1细胞生长
3 讨论
EYA蛋白因其C端有271个氨基酸残基构成的保守性较高的EYA结构域而得名,这些蛋白普遍存在于从果蝇到人类的各种生物体中[3],在体内发挥多种作用。作为重要的转录调控因子,它们参与EGFR/RAS/MAP、TGFβ/DPP、Wingless、Hedghog 和Notch[4-8]等信号通路,在胚胎发育过程中调控细胞增殖、组织分化和器官如性腺、肌、四肢、神经、腺和肾脏等的发育[9-11],EYA基因发生突变或表达异常将导致鳃-耳-肾综合征、鳃裂-耳综合征等遗传病。同时,EYA蛋白是一类蛋白磷酸酯酶,可去磷酸化酪氨酸磷酸化抗原和苏氨酸/丝氨酸磷酸化抗原。各种原因导致细胞内DNA双键断裂后,EYA3即可逐渐去磷酸化DNA修复关键蛋白——组蛋白H2A变体H2AX的142位的酪氨酸残基[12]。研究还表明EYA蛋白在多种癌症中发挥作用:在卵巢癌、恶性外周神经鞘瘤中,EYA蛋白的过表达与肿瘤生长和转移相关[13];EYA3在尤文肉瘤中高表达,敲低EYA3能降低尤文肉瘤细胞的存活率,且使细胞对DNA损伤的化疗敏感[14];在浸润性乳腺癌细胞系MDA-MB-231中沉默EYA3的表达能抑制细胞的转移[15]。
综上所述,EYA3是一个与癌症有密切关系的多功能癌基因,我们构建的EYA3基因的真核表达载体,为进一步研究其在癌症发生发展中的分子生物学机制奠定了基础。
[1]Silver S J,Davies E L,Doyon L,et al.Functional dissection of eyes absent reveals new modes of regulation with in the retinal determination gene network[J].Mol Cell Biol,2003,23(17):5989-5999.
[2]Xu P X,Cheng J,Epstein J A,et al.Mouse Eya genes are expressed during limb tendon development and encode a tran⁃scriptionalactivation function[J].Proc NatlAcad SciUSA,1997,94(22):11974-11979.
[3]Epstein J A,Neel B G.Signal transduction:an eye on organ development[J].Nature,2003,426(6964):238-239.
[4]Hsiao F C,Williams A,Davies E L,et al.Eyes absent medi⁃ates cross-talk between retinal determination genes and the re⁃ceptor tyrosinekinase signaling pathway[J].Dev Cell,2001,1(1):51-61.
[5]Baonza A,Freeman M.Control of Drosophila eye specification by Wing lesssignaling[J].Development,2002,129(23):5313-5322.
[6]Bai J,Montell D.Eyes absent,a key repressor of polar cell fate during Drosophila oogenesis[J].Development,2002,129(23):5377-5388.
[7]Kumar J P,Moses K.EGF receptor and Notch signaling act upstream of Eyeless/Pax6 to control eye specification[J].Cell.2001,104(5):687-697.
[8]Kenyon K L,Ranade S S,Curtiss J,et al.Coordinating prolif⁃eration and tissue specification to promote reional identity in the Drosophila head[J].Dev Cell,2003,5(3):403-414.
[9]Fabrizio J J,Boyle M,DiNrdo S.A somatic role for eyes ab⁃sent(eya)and sine oculis(so)in Drosophila spermatocyte devel⁃opment[J].Dev Biol,2003,258(1):117-128.
[10]Li X,Perissi V,Liu F,et al.Tissue-specific regulation of ret⁃inal and pituitary precursor cell proliferation[J].Science,2002,297(5584):1180-1183.
[11]Xu P X,Zheng W,Laclef C,et al.Eya1 is required for the morphogenes is of mammalian thymus,parathyroid and thy⁃roid[J].Development,2002,129(13):3033-3044.
[12]Singh N,Basnet H,Wiltshire T D,et al.Dual recognition of phosphoserine and phosphotyrosine in histone variant H2A.X by DNA damage response protein MCPH1[J].Proc Natl Acad Sci USA,2012,109(36):14381-14386.
[13]Tadjuidje E,Wang T S,Pandey R N,et al.The EYA tyro⁃sine phosphatase activity is pro-angiogenic and is inhibited by benzbromarone[J].PLoS One,2012,7(4):e34806.
[14]Robin T P,Smith A,McKinsey E,et al.EWS/FLI1 regulates EYA3 in Ewing sarcoma via modulation of miRNA-708,re⁃sulting in increased cell survival and chemoresistance[J].Mol Cancer Res,2012,10(8):1098-1108.
[15]Pandey R N,Rani R,Yeo E J,et al.The Eyes Absent phos⁃phatase-transactivator proteins promote proliferation,transforma⁃tion,migration,and invasion of tumor cells[J].Oncogene,2010,29(25):3715-3722.
猜你喜欢
环球时报(2022-09-20)2022-09-20
传染病信息(2021年6期)2021-02-12
今日农业(2020年24期)2020-12-15
科海故事博览·下旬刊(2019年6期)2019-04-16
食品科学(2018年10期)2018-05-23
兽医导刊(2016年12期)2016-05-17
西南医科大学学报(2015年1期)2015-08-22
中国当代医药(2015年9期)2015-03-01
生物医学工程学进展(2015年1期)2015-02-28
化学工业与工程(2015年1期)2015-02-10
