
miR-30a和miR-30b靶向GW182的生物学功能研究
2013-10-29 09:36苏雪婷夏伟陈颖李少华丁红梅李慧黄皑雪赵强李洁邵宁生
生物技术通讯 2013年5期
苏雪婷,夏伟,陈颖,李少华,丁红梅,李慧,黄皑雪,赵强,李洁,邵宁生
军事医学科学院 基础医学研究所,北京 100850
microRNA(miRNA)是近年发现的一类高度保守、内源性非蛋白编码的20~25个核苷酸的小分子RNA,主要在转录后水平调节基因的表达。miRNA经典的加工成熟途径是:首先细胞核内编码miRNA的基因通过RNA聚合酶转录成长的初级miRNA(pri-miRNA)[1],接 着 pri-miRNA 在 一 种 RNaseⅢ(Drosha酶)及其伴侣分子(DGCR8)组成的复合物作用下被剪切为70~90个核苷酸长度、具有茎环结构的前体 miRNA(pre-miRNA),pre-miRNA 在 Ran-GTP依赖的核质P细胞质转运蛋白Exportin-5的作用下从核内运输到胞质中[2],随后pre-miRNA在另一种RNaseⅢ(Dicer酶)的作用下被剪切成包含21~25个核苷酸,且5'端磷酸化、3'端有2个核苷酸的突起的类似于siRNA的不完全配对的双链RNA,双链RNA是由成熟miRNA与miRNA组成的二聚体,miRNA与miRNA位置相互对应,最后在RNA解旋酶作用下生成单链miRNA,成熟miRNA结合到RNA诱导的基因沉默复合物(RNA-induced silencing complex,RISC)[3]中发挥作用,成熟miRNA通过与靶基因3'UTR的结合抑制翻译或直接降解mRNA[4-7]。近期研究显示,RISC翻译抑制机制是由RISC捕获靶mRNA,引导mRNA进入细胞质中的某些特殊结构,最终使得mRNA逃逸翻译程序。人们把细胞质中的这些特殊结构命名为P-body(processing bod⁃ies)[8]。GW182是P-body的标志性分子,对维持P-body的功能与完整性至关重要,因此P-body通常又被称为GW-body[9]。尽管许多研究已证实GW/P-body在转录后基因表达调控过程中发挥重要作用,但由于其组成结构与功能十分复杂,人们对它的理解仍不足以完全解释其形成机制与生物学意义。
miR-30家族包括miR-30a、miR-30b、miR-30c、miR-30d和miR-30e,作为miRNA的重要组成部分,在人和动物miRNA的功能研究中起着不容忽视的作用。目前对miR-30a和miR-30b的研究报道仍较少。有研究表明,miR-30家族能通过靶向调节线粒体分裂来调节细胞凋亡[10]。miR-30b还与精神分裂症和成神经管细胞瘤[11]等有一定的关系。
我们通过生物信息学预测并确定miR-30a和miR-30b能够直接靶向GW182,并通过下调GW182的表达影响其他miRNA/siRNA对自身靶基因的作用,揭示了miR-30a、miR-30b调控miRNA/siRNA生物学途径的新功能。
1 材料与方法
1.1 材料
人宫颈癌细胞系HeLa细胞和大肠杆菌DH5α由本室保存;萤光素酶报告基因载体pGL3-Control及海肾萤光素酶载体pRL-CMV购自Promega公司,由本室保存;绿色荧光蛋白载体(pEGFP-N2)购自In⁃vitrogen公司;细胞培养用DMEM培养基购自Gibco公司;胎牛血清购自Hyclone公司;总RNA提取所用TRIzol试剂购自Sigma公司;M-MLV反转录酶、萤光素酶检测试剂盒、实时定量荧光PCR试剂盒、限制性内切酶XbaⅠ和NdeⅠ、T4DNA连接酶购自Promega公司;阳离子脂质体转染试剂LipofectAMINE 2000购自Invitrogen公司;兔单抗GW182购自Santa Cruz公司;兔单抗GAPDH购自北京康维世纪有限公司;BCA蛋白定量试剂盒为碧云天生物技术有限公司产品;超敏发光液Pico购自Thermo公司;miR-30a、miR-30b mimics,GW182 siRNA,NC(阴性对照)为广州瑞博公司产品;其他相关试剂均为进口分装或国产分析纯产品;引物由上海生工生物工程公司合成。
1.2 细胞培养与细胞转染
HeLa细胞在DMEM(含5%胎牛血清)培养基中,于37℃、5%CO2、饱和湿度条件下培养。将HeLa细胞按2×105/mL的密度种于细胞培养板中,培养24 h后留待转染。每孔需萤光素酶报告载体100 ng,海肾萤光素酶载体 5 ng,miR-30a、miR-30b mimics或 NC 20 pmol,稀释 于 50 μL 无 血 清DMEM培养基中,轻弹混匀;按照样品与转染试剂为1∶3的比例将 LipofectAMINE2000转染试剂 1.5 μL稀释于50 μL无血清DMEM培养基中,轻弹混匀,室温静置5 min;将稀释好的转染试剂加入载体稀释物中,轻弹混匀,室温静置20 min;将样品-转染试剂混合物逐滴加入细胞培养板中,前后晃动3~5次,培养4 h后更换培养基,继续培养48 h后检测荧光。
1.3 miRNA靶基因预测
预测数据库为TargetScan 5.2版本。
1.4 双萤光酶素报告基因检测
1.4.1 miRNA重组萤光素酶报告载体的构建 根据GW182的3'UTR序列,在上下游分别插入XbaⅠ、NdeⅠ酶切位点,从HeLa细胞基因组DNA中扩增包含miR-30a、miR-30b 4个保守靶位点的3条基因片段。将PCR钓取的包含miR-30a、miR-30b靶位点的片段连接至萤光素酶报告载体pGL3-Control中,鉴定出的阳性克隆送上海生工生物工程公司测序。
1.4.2 荧光检测 转染48 h后的细胞培养板用1×PBS缓冲液洗2次,每孔加入100 μL裂解缓冲液,室温裂解10 min;收集细胞裂解物,室温5000 r/min离心 5 min,吸取上清待测;取 50 μL上清,加入 Lucif⁃erase Assay ReagentⅡ(LAR液)50 μL,轻轻混匀后检测荧光值;取出上清-LAR反应液,再加入Stop&Glo Reagent(S液)50 μL,轻弹混匀后检测荧光值;计算2次检测结果的比值,对每组间的比值进行比较。
1.5 Western印迹
转染48 h后的细胞用冰冷的1×PBS洗2次,冰上将细胞用刮刀刮下,低速离心收集细胞,加入60 μL预冷的RIPA裂解缓冲液,冰上裂解30 min后,4℃、12 000 r/min离心30 min,收集上清;BCA法蛋白定量;10%SDS-PAGE凝胶电泳分离蛋白,湿转至NC膜;用含5%脱脂奶粉及0.1%吐温-20的TBST缓冲液室温封闭1 h;根据抗体说明书室温孵育一抗1 h(稀释比例1∶100),TBST缓冲液洗4次,每次10 min;室温孵育山羊抗兔辣根过氧化物酶标记的IgG(稀释比例1∶8000)1 h,洗膜6次,ECL显色。
1.6 RNA提取与实时荧光定量PCR
转染48 h后的细胞用1×PBS洗2次,每孔加入1 mL TRIzol试剂,室温裂解至细胞裂解物清亮;加入 200 μL 氯仿,剧烈振荡 10 s,室温静置 5 min;4℃、12 000 r/min离心15 min;小心吸取上层水相移至新管,加入500 μL异丙醇,颠倒混匀后室温静置30 min;4℃、12 000 r/min离心15 min后可见管底白色沉淀即为总RNA;吸弃上清,加入1 mL 80%乙醇颠倒混匀;4℃、10 000 r/min离心15 min;弃上清,室温晾干沉淀,DEPC水溶解;取适量RNA样品稀释,紫外分光光度计测定浓度,读取D260nm/D280nm比值测定纯度,电泳鉴定RNA完整性;取提取的总RNA 1 μg作为模板,用M-MLV反转录酶反转录合成cDNA;以cDNA为模板,用Stratagene 3000p实时荧光定量PCR系统及Gotaq实时荧光定量试剂盒检测GW182 mRNA的表达量[引物:GW182 F(CTCT GTGGATGCTCCTGAAAG)和 GW182 R(TGCTTGG ATTTAACCCTCCATTT)]。
2 结果
2.1 miR-30a、miR-30b靶向GW182
2.1.1 生物信息学预测 采用生物信息学数据库TargetScan 5.2预测分析,GW182的3'UTR序列包含miR-30a、miR-30b的4个保守靶位点(图1)。
2.1.2 GW182 3'UTR miR-30a、miR-30b靶位点钓取 以HeLa细胞基因组DNA为模板,获取包含miR-30a、miR-30b的4个靶位点的GW182 3'UTR的3段片段S1、S2和S3(图2)。
2.1.3 重组萤光素酶报告载体的构建及荧光检测将PCR扩增得到的片段插入萤光素酶报告载体中,挑取阳性克隆测序。将插入了GW182 3'UTR片段的萤光素酶载体分别与miR-30a、miR-30b、NC共转染HeLa细胞,48 h后收集细胞,检测萤光素酶活性。结果显示,转染了miR-30a、miR-30b的细胞与同时转染了NC的细胞相比,每个靶位点的萤光素酶活性均下降了20%以上,其中S2、S3相比S1具有更显著差异。结果说明miR-30a、miR-30b能够直接靶向GW182 3'UTR(图3)。
2.2 miR-30a、miR-30b靶向GW182的生物学功能
2.2.1 转染miR-30a、miR-30b后GW182 mRNA的表达变化 将 GW182 siRNA、miR-30a、miR-30bmimics及NC分别转染HeLa细胞,48 h后收集细胞提取总RNA进行反转录,通过荧光定量PCR检测转染后GW182 mRNA的水平。与NC组相比,转染了GW182 siRNA、miR-30a、miR-30b mimics的实验组中GW182的表达水平下降了40%以上。miR-30a、miR-30b mimics能够明显抑制GW182在mRNA水平上的表达(图4)。
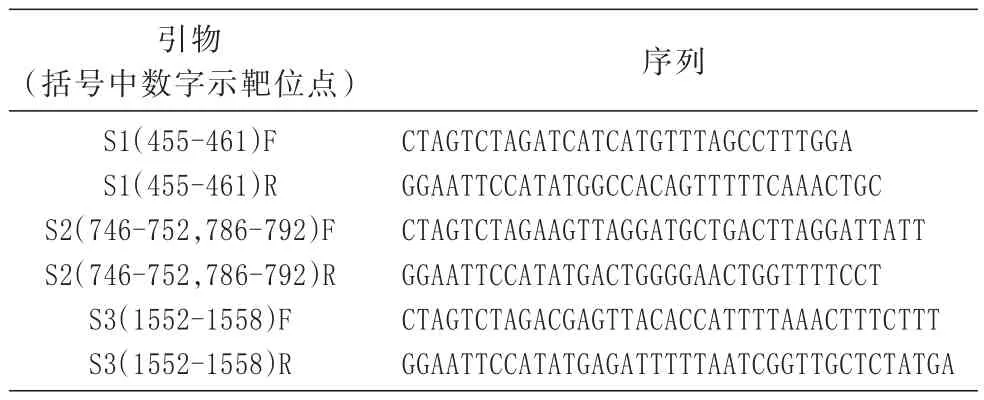
表1 钓取靶位点所用引物及序列
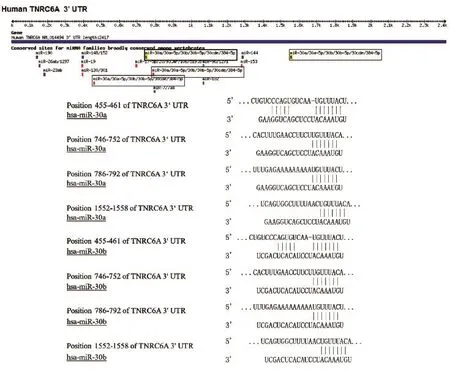
图1 TargetScan 5.2预测GW182的3'UTR区miR-30a、miR-30b靶位点
2.2.2 转染miR-30a、miR-30b后GW182蛋白的表达 变 化 将 GW182siRNA、miR-30a、miR-30b mimics及NC分别转染HeLa细胞,48 h后收集细胞提取总蛋白,通过Western印迹检测转染后GW182的蛋白表达水平。与NC组相比,转染了GW182 siRNA、miR-30a、miR-30b mimics的实验组GW182蛋白表达水平明显下降。miR-30a mimic能够抑制GW182在蛋白水平上的表达(图5)。

图2 GW182 3'UTR片段扩增
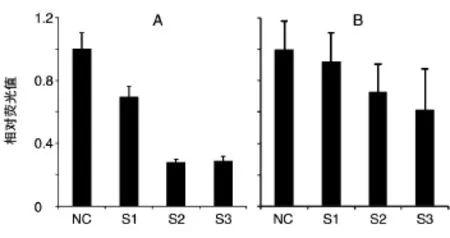
图3 转染miR-30a、miR-30b后的萤光素酶活性
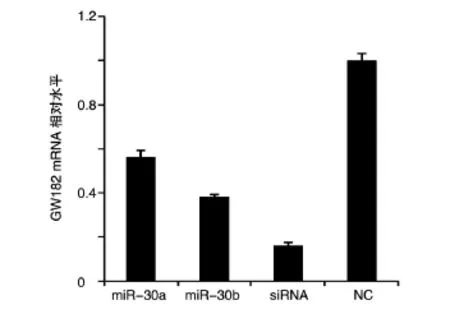
图4 转染GW182 siRNA、miR-30a、miR-30b mimics及NC后GW182 mRNA的表达变化
2.3 miR-30a、miR-30b影 响mir-200b对 靶 基 因ZEB1的调控
GW182是P-body的标志性分子,抑制GW182的表达将影响miRNA作用的生物学途径。我们用文献报道的miR-200b直接靶向ZEB1[12]来验证miR-30a、miR-30b对GW182生物学功能的调控。在 24孔板中分别转染 miR-30a、miR-30b、GW182 siRNA及NC,24 h后同时转染miR-200b及NC,48 h后收集细胞总RNA及总蛋白,利用荧光定量PCR及Western印迹检测转染后ZEB1的mRNA和蛋白变化。结果显示,miR-200b靶向ZEB1,转染miR-200b后 ZEB1被下调。利用 miR-30a、miR-30b、GW182 siRNA将细胞内GW182蛋白水平下调后再转染miR-200b,结果显示miR-200b对ZEB1的表达抑制作用显著下调。这说明HeLa细胞内转染miR-30a、miR-30b后通过靶向GW182而影响了mir-200b对靶基因ZEB1的调控(图6)。
2.4 转染miR-30a、miR-30b后影响GFP siRNA对GFP表达的调控
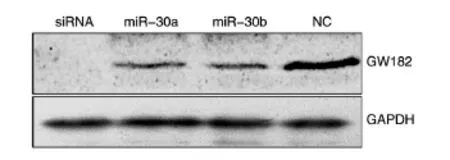
图5 转染GW182 siRNA、miR-30a、miR-30b mimics及NC后GW182蛋白水平的表达变化
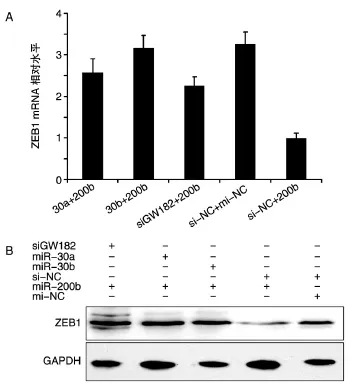
图6 HeLa细胞内转染miR-30a、miR-30b后影响mir-200b对靶基因ZEB1的调控
为了探索体内转染miR-30a、miR-30b对siRNA沉默效应的影响,我们在24孔板中分别转染miR-30a、miR-30b、GW182 siRNA及NC,24 h后同时转染 pEGFP-N2及 GFP siRNA,24~48 h后在荧光显微镜下观察GFP的表达情况。结果显示,pEGFPN2与GFP siRNA共转染时,GFP的表达被明显抑制,当先转染miR-30a、miR-30b、GW182 siRNA 将细胞内GW182蛋白下调后,再转染pEGFP-N2与GFP siRNA,GFP siRNA对GFP的沉默作用明显受到抑制(图7)。说明HeLa细胞中miR-30a、miR-30b通过靶向GW182能够减弱siRNA对靶基因的沉默。
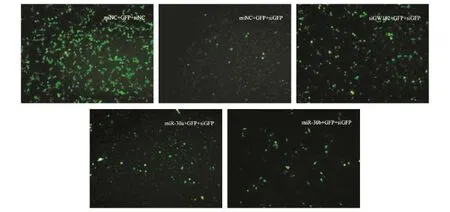
图7 HeLa细胞内转染miR-30a、miR-30b后影响GFP siRNA抑制pEGFP-N2的表达
3 讨论
很长时间以来,miRNA/siRNA针对靶基因的抑制效应已成为调控生物基因组表达的公认的决定性因素之一。这些小的非编码RNA依靠转录后调控途径参与着调控生物体生长发育、细胞凋亡、神经分化和免疫等重要过程。Dicer酶、Argonaute蛋白家族等传统明星蛋白分子在转录后调控过程中发挥的功能已有很多报道。GW182蛋白作为GW/P-body的重要组成部分,其结构与功能的重要性也逐渐引起广泛关注。1997年Bashkiron等[13]首次报道了一种在哺乳动物细胞质的特定区域富集的核酸外切酶XRN1;2002年Eystathioy等[9]又一次发现了这种相对分子质量为182×103、包含Gly和Tyr重复序列的蛋白,并将它命名为GW182。随着研究的逐渐展开,GW/P-body的多种组分不断被鉴定,其中包括大量蛋白质分子。同时,GW182也能与细胞质中的poly(A)结合蛋白(PABP)、PAN2-PAN3及CCR4-NOT脱腺苷化酶复合物相互作用组成蛋白复合体[14]。这一系列发现表明,GW182的功能是作为组成多蛋白复合体的骨架蛋白,而这种蛋白复合物组装完毕后则发挥它们沉默miRNA靶标的作用。最近有文献报道,稳定敲除GW182或它的同源蛋白TNRC6B,会缩减转染后miRNA mimic的半衰期;如果恢复GW182的表达,则会显著增强转染后miRNA mimic的稳定性[15]。基于此,我们分别利用生物信息学预测、双萤光素酶报告载体系统、实时荧光定量PCR、Western印迹等实验手段发现并证明了miR-30a、miR-30b能够靶向GW182。并且,在HeLa细胞中通过转染miR-30a、miR-30b mimics与GW182 siRNA不同程度的下调GW182后,影响下游miR-200b对其靶基因ZEB1的调控,以及GFP siRNA对GFP表达的抑制。这些结果揭示了miR-30a、miR-30b通过靶向GW182影响siRNA/miRNA作用途径的新功能。通过靶向miRNA/siRNA生物学途径中的关键作用分子而影响miRNA/siRNA自身加工过程或功能效应的一系列miRNA将会越来越受重视。
[1]Lee Y,Kim M,Han J,et al.MicroRNA genes are tran⁃scribed by RNA polymerase II[J].EMBO J,2004,23:4051-4060.
[2]Lund E,Güttinger S,Calado A,et al.Nuclear export of mi⁃croRNA precursors[J].Science,2004,303(5654):95-98.
[3]Hammond S M,Bernstein E,Beach D,et al.An RNA direct⁃ed nuclease mediatespost-transcriptionalgene silencing in Drosophila cells[J].Nature,2000,404(6775):293-296.
[4]Newman M, Thomason J, Hammond S. Lin-28 interaction with the let-7 precursor loop mediates regulated microRNA processing[J].RNA,2008,14(8):1539-1549.
[5]Rycak A,Fuchs H,Smirnova L,et al.A feedback loop com⁃prising lin-28 and let-7 controls pre-let-7 maturation during neural stem-cell commitment[J].Cell,2008,10(8):987-993.
[6]Viswanathan S R,Daley G Q,Gregory R I.Selective block⁃ade of microRNA processing by Lin 28[J].Science,2008,320(5872):97-100.
[7]Lytle J R,Yario T A,Steitz A.Target m RNAs are re⁃pressed as efficiently by microRNA-binding sites in the 5'UTR as in the 3'UTR[J].Proc Natl Acad Sci USA,2007,104(23):9667-9672.
[8]Eulalio A,Behm-Ansmant I,Izaurralde E.P bodies:at the crossroadsofpost-transcriptionalpathways[J].NatRev Mol Cell Biol,2007,8(1):9-22.
[9]Eystathioy T,Chan E K,Tenenbaum S A,et al.Aphosphory⁃lated cytoplasmicautoantigen,GW182,associates with aunique population of human mRNAs within novel cytoplasmicspeckles[J].Mol Biol Cell,2002,13(4):1338-1351.
[10]Li J,Donath S,Li Y,et al.miR-30 regulates mitochondrial fission through targeting p53 and the dynamin-related pro⁃tein-1 pathway[J].PLoS Genet,2010,6(1):e1000795.
[11]Quintavalle C,Donnarumma E,IaboniM,etal.Effectof miR-21 and miR-30b/c on TRAIL-induced apoptosis in glio⁃ma cells[J].Oncogene,2012,doi:10.1038/onc.2012.410.
[12]Gregory P A,Bert A G,Paterson E L,et al.The miR-200 family and miR-205 regulate epithelial to mesenchymal transi⁃tion by targeting ZEB1 and SIP1[J].Nat Cell Biol,2008,10:593-601.
[13]Bashkirov V I,Scherthan H,Solinger J A,et al.Amouse cyto⁃plasmic exoribonuclease(mXRN1p)with preference for G4 tetra⁃plex substrates[J].J Cell Biol,1997,136(4):761-773.
[14]Braun J E,Huntzinger E,Izaurralde E.The role of GW182 proteins in miRNA-mediated gene silencing[J].Adv Exp Med Biol,2013,768:147-163.
[15]Yao Bing,La L B,Chen Ying-Chi,et al.Defining a new roleofGW182 in maintaining miRNA stability[J].EMBO Rep,2012,13(12):1102-1108.
猜你喜欢
煤气与热力(2021年2期)2021-03-19
传奇故事(破茧成蝶)(2018年6期)2018-11-14
凤凰生活(2018年8期)2018-09-03
传奇故事(上旬)(2018年6期)2018-07-11
作文与考试·小学高年级版(2017年13期)2017-07-12
中国水稻科学(2016年5期)2016-10-25
物理化学学报(2015年7期)2015-12-30
橡胶工业(2015年7期)2015-08-29
橡胶工业(2015年8期)2015-07-29
中国药业(2014年21期)2014-05-26
