
美洲商陆光合光响应曲线及其模型拟合
2013-10-26 05:49:00闫小红胡文海
井冈山大学学报(自然科学版) 2013年3期
闫小红,胡文海,王 宁,邹 谭
美洲商陆光合光响应曲线及其模型拟合
*闫小红1,2,胡文海1,王 宁1,邹 谭1
(1. 井冈山大学生命科学学院,江西,吉安 343009;2. 南京林业大学森林资源与环境学院,江苏,南京 210037)
为探讨不同光响应模型对美洲商陆光合特性的适用性,利用4种典型的光响应模型拟合了美洲商陆的光响应曲线,通过分析光合参数模拟值与实测值的拟合相似度,在4个模型中筛选最佳美洲商陆光响应模型。结果表明,直角双曲线模型、非直角双曲线模型、指数方程以及修正的直角双曲线模型对美洲商陆的光合光响应曲线都可以进行拟合,决定系数均为2>0.99,但只有修正的直角双曲线模型可以计算它们的饱和光强。由各模型计算的光合参数可知,非直角双曲线模型求得的n max远高于实测值,指数方程与修正的直角双曲线模型求得的n max与实测值最为接近。直角双曲线、非直角双曲线模型以及指数方程求得的sat均远低于实测值,唯有修正的直角双曲线模型求得的sat与实测值最为相符。因此,修正的直角双曲线模型对于美洲商陆的光合光响应曲线的拟合以及各光合参数的求取最为适合。由修正的直角双曲线模型计算得到的美洲商陆的光合参数可知,美洲商陆的n max为16.98 μmol·m-2·s-1,sat为1614.54 μmol·m-2·s-1,c为14.14 μmol·m-2·s-1,表明美洲商陆对光强具有较大的适应范围。
美洲商陆;光响应曲线;直角双曲线修正模型;饱和光强
0 引言
植物光合作用对光响应模型(或光响应曲线)研究的是植物净光合速率和光合有效辐射之间的关系,对了解植物光化学过程中的光化学效率非常重要[1]。还可由光合作用对光响应曲线估算植物的饱和光强、最大净光合速率、光补偿点、暗呼吸速率和表观量子效率等重要的光合参数[2]。其中,饱和光强反映了植物利用光强的能力,其值高说明植物在受到强光时生长发育不易受到抑制;叶片的最大净光合速率反映了植物叶片的最大光合能力;光补偿点反映的是植物叶片光合作用过程中光合同化作用与呼吸消耗相当时的光强;表观量子效率反映了植物在弱光情况下的光合能力。
目前,用于拟合植物光合光响应曲线的模型很多,最常用的模型有直角双曲线模型[3-4]、非直角双曲线模型[5-10]、指数方程[11-12]和直角双曲线的修正模型[13-14]等。但不同的植物有其相适应的光合光响应曲线模型,如指数方程,用于拟合藻类、海藻、海草及其他浮叶植物或沉水植物的光响应曲线的较多[15-16],也有用于拟合高等植物的[17]。而目前最常用的模型为非直角双曲线模型[18-19]。随着直角双曲线的修正模型的系数的生物学意义的明确[20]及对绝大多数植物光响应曲线拟合的适应性,此模型逐渐得到推广应用[21-22]。只有采用正确的光合光响应曲线的模型才能计算得到准确的最大净光合速率(n max)、饱和光强(sat)、光补偿点(c)及表观量子效率()等衡量一种植物光合能力的重要的光合参数,才可能为后续的研究(如饱和光强下的CO2响应曲线等)提供可靠的数据。
因此,我们在研究美洲商陆光合特性的同时,采用不同模型拟合美洲商陆的光响应曲线,得到最适于美洲商陆光合光响应曲线的拟合模型,并判断这些模型的优势以及潜在问题,为进一步推动植物光合作用对光响应在植物生理生态研究中的应用提供理论基础。同时也为后续美洲商陆实验的开展提供理论与实践依据。
1 材料与方法
1.1 供试材料
于2012年5月中旬,取一年生株高为15 cm左右的美洲商陆幼苗定植于直径30 cm的塑料花盆中,置于温室中培养,待株高为25 cm左右,叶片数为7~8片叶时,置于室外,适应1周,每株挑选自上而下第2~4片叶进行测量,4次重复。
1.2 光响应数据的测定
自然光诱导1~1.5h后,采用LI-6400XT便携式光合测定系统进行测定,CO2浓度控制在(400 ± 2) μmol·mol-1,温度控制在(30 ± 2) ℃,空气相对湿度为50% ~ 70%,光合有效辐射的梯度设置为1500、1200、1000、800、600、400、200、150、100、50、20和0 μmol·m-2·s-1下测定美洲商陆的净光合速率(n)(μmol·m-2·s-1)、气孔导度和胞间CO2浓度等,然后用各模型拟合光响应曲线,得到光响应曲线的初始斜率(α)、饱和光强(sat)、最大净光合速率(n max)、光补偿点(c)和暗呼吸速率(d)。
1.3 光响应曲线模型
1.3.1 直角双曲线模型
直角双曲线模型[3]的数学表达式为:
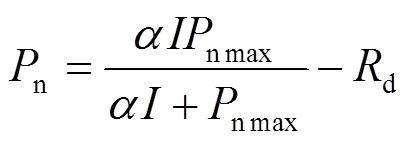
式中,n为净光合速率,为光强,α为植物光合作用对光响应曲线在= 0时的斜率,即光响应曲线的初始斜率,也称为初始量子效率,n max为最大净光合速率,d为暗呼吸速率。式(1)是一个没有极值的函数,也即直角双曲线是一条没有极点的渐近线。因此,无法由(1)式直接求出植物的n max和饱和光强,即无法用(1)式求解植物饱和光强的解析解。为了估算植物的饱和光强,就需要用直线方程拟合弱光强条件下(≤200 μmol·m-2·s-1)的光响应数据得到表观量子效率(apparent quantum efficiency,),利用非线性最小二乘法估算n max,而对于sat,需要求解直线方程式(2)得到[2]:
n max=×sat-d(2)
1.3.2 非直角双曲线模型
光响应曲线的非直角双曲线模型[5]的表达式为:
(3)
式中:n、、、n max、d的定义同前,为反映光响应曲线弯曲程度的曲角参数,取值0 <≤1。式(3)同样是一个没有极值的函数,无法求取sat的解析解,估算sat的方法与直角双曲线模型的方法相同。
1.3.3 指数函数模型
光响应曲线的指数函数模型形式较多,由Bassman 和Zwier 在1991年首次给出,随后Prado 和Moraes 1997年、Waltling等在2000年也提出过相关模型。但它们都是没有极值的函数。本文采用Brassman 和 Zwier 提出的指数函数模型[11],其表达式为:

式中:n、、、n max、d的定义同前,e 为自然对数的底。通过式(4)可以求取c的解析解,但无法求取sat,要估算饱和光强,需假设光合速率为0.9n max或0.99n max所对应的光强为饱和光强[23-24]。
1.3.4 直角双曲线的修正模型
直角双曲线的修正模型是Ye 和Yu提出的[13-14],其表达式为:

式中:、、d的定义同前,称为光抑制项和称为光饱和项[20]。由于式(5)是一个具有极值的函数,因此,由式(5)可以直接求取sat、c和n max的解析解。同时,可以拟合具有光抑制的光响应曲线。
1.4 数据处理和分析
实验数据处理及作图由Excel 2007完成,模型分析由SPSS 11.5软件分析。
2 结果与分析
2.1 美洲商陆光响应曲线的拟合及其比较
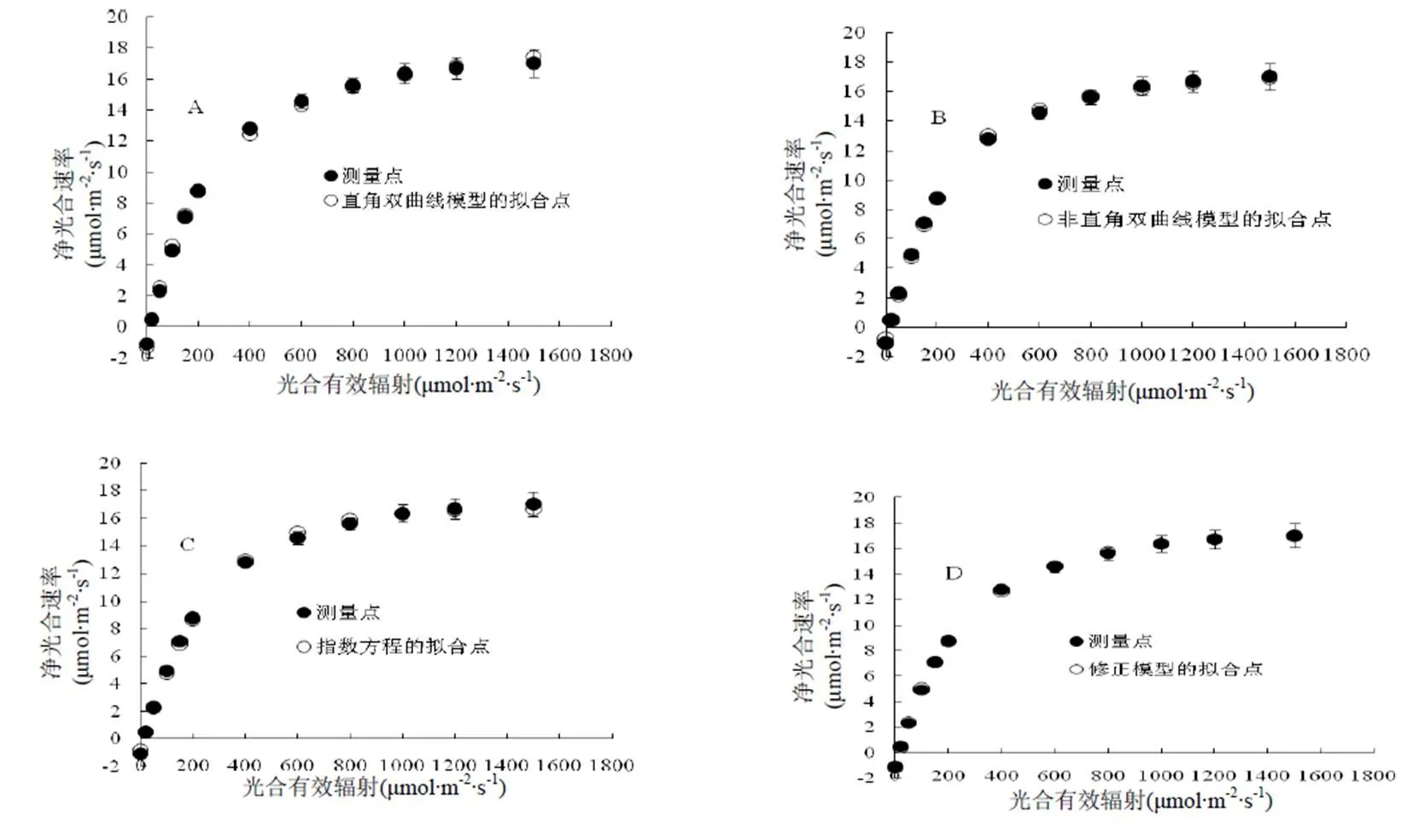
图1 4种模型拟合的美洲商陆的光响应曲线
图1A,B,C,D为美洲商陆光合作用光响应曲线测量点与4种模型的拟合点。由图1可知,净光合速率随光合有效辐射的增加而增加,接近饱和光强时增加缓慢。4种模型在各个数据点下的拟合值与实测值都较为接近,以修正的直角双曲线模型各数据点下的拟合值与实测值最为接近,测量点与拟合点基本是完全重合的(图1 D)。因此,对于拟合美洲商陆的光响应曲线的模型来说,4种模型均适用,但以修正的直角双曲线模型最为适合。
2.2 美洲商陆的光响应曲线参数比较
表1给出了4种光响应曲线模型拟合美洲商陆的光响应曲线所得到的相关参数及相应的测量值。由表1可知,各模型拟合下的初始量子效率(α)均在0 < α ≤ 0.125范围之内,但非直角双曲线模型拟合下的α已经达到了它的最大值0.125。由非直角双曲线模型拟合的美洲商陆的最大净光合速率(n max)远高于实测值,为实测值的2.23倍。其余3种模型拟合的最大净光合速率与实测值较为接近,以修正的直角双曲线模型拟合效果最好。对于光饱和点(sat),由直角双曲线、非直角双曲线模型通过直线方程求解得到的sat远低于实测值,分别低于实测值1032.60 μmol·m-2·s-1和706.37 μmol·m-2·s-1。而指数方程为假设n为0.9n max所对应的光强sat,其值也明显低于实测值,相差740.81 μmol·m-2·s-1,如果按0.99n max进行假设则无法求取sat。而由修正的直角双曲线模型计算得到的sat与实测值最为接近,仅相差114.54 μmol·m-2·s-1。4种模型拟合下得到的光补偿点(c),除了非直角双曲线模型远低于实测值外,其余3种模型之间相差不大,略低于实测值。4种模型拟合下得到的暗呼吸速率d与实测值较为接近,说明4种模型对于光响应曲线低光强部分的拟合效果均较好。
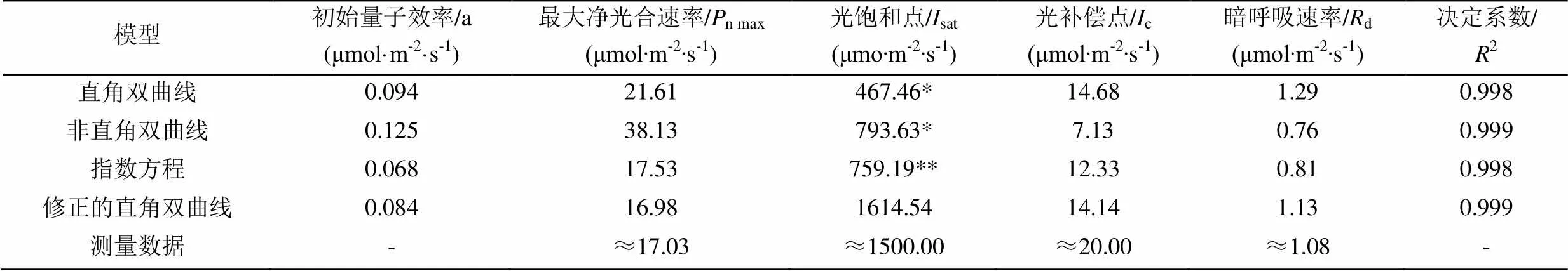
表1 4种光响应曲线模型拟合美洲商陆光响应曲线的实测值与拟合结果
*,用方程n max=×sat-d求得的光饱和点;**,假定净光合速率为0.9n max所对应的光强为光饱和点。
由表1可知,尽管直角双曲线、非直角双曲线、指数方程求得的决定系数2>0.99,但它们拟合得到的光合参数与实测值相差较大,因此,正如图1中所显示的,2越大只能说明模型拟合程度越高,并不能保证拟合结果一定与实测值相符。由表1中各光合参数可知,修正的直角双曲线模型求解得到的各项光合参数与实测值最为吻合。因此,采用修正的直角双曲线模型对于拟合美洲商陆光合作用光响应曲线最为适宜。
2.3 美洲商陆的光合特性
由表1可知,美洲商陆在修正的直角双曲线模型的拟合下,其光补偿点为(14.14 μmol·m-2·s-1),其光补偿点较低,说明其在较弱的光环境下,也能够积累一定的光合产物。其饱和光强为1614.54 μmol·m-2·s-1,也表现了较高的饱和光强,说明美洲商陆对光照的适应范围较宽,对光照强弱的耐受能力较强,但其并没有表现出较高的最大净光合速率(16.98 μmol·m-2·s-1)。
3 讨论
已有很多研究表明,直角双曲线模型和非直角双曲线模型在实际应用过程中,会出现饱和光强远低于实际测量值,而最大光合速率远大于实际测量值[25-28],本实验中对于美洲商陆的光合光响应曲线的模型拟合也出现了上述问题,说明上述两个模型虽然可以很好的拟合绝大多数植物的光响应曲线,但其计算得到的光合参数与实测值相差很大,并且无法处理光抑制条件下的光响应数据[25-28]。
指数方程也是用于光响应曲线的主要拟合方程,主要用于拟合藻类、海草和其他沉水植物的光响应曲线,但对于本文中美洲商陆用该方程拟合的光响应曲线,其求得的n max比直角双曲线与非直角双曲线求得的n max更为接近实测值,但求得的sat,则与实测值相差很大,且它是假定0.9n max时的光强为饱和光强,如果假定0.95n max或0.99n max时,则用指数方程不能得到美洲商陆的饱和光强。因此,用指数方程计算的饱和光强具有人为性,与实际不相符。
而修正的直角双曲线模型[13-14]克服了其它几个模型拟合光响应曲线光合参数时出现的上述各种问题,不但能直接求取最大净光合速率、饱和光强等参数的解析解,且得到的光合参数与实测值非常接近。因此,建议在应用数学模型拟合植物的光响应曲线时,采用修正的直角双曲线模型。
此外,由修正直角双曲线模型参数的生物学意义可知[20],β为光抑制项,γ值一定时,β值越大,光响应曲线弯曲程度就变大,表示PSⅡ天线色素分子吸收光量子的截面与PSⅡ天线色素分子处于激发态的平均寿命的乘积越大,植物受光抑制就越强。而γ为光饱和项,当β一定时,γ值越小,表示光响应曲线弯曲程度就越小,植物就越不容易发生光饱和现象,对应饱和光强的值就越大。因此,修正的直角双曲线模型不仅可以很好的拟合植物的光响应曲线,且其参数β与γ具有生物学意义,也体现了此模型相对于其他模型的优点及更广泛的适应性。
美洲商陆具有抑菌、抗病、重金属积累等方面的作用,对其光合特性的研究,有助于探明其在上述几个方面,尤其是重金属积累方面的光合生理机制,具有重要的实际意义。本实验以温室内盆栽的美洲商陆作为实验材料,研究美洲商陆的光合特性。研究发现美洲商陆具有较高的饱和光强,野外观察也发现,美洲商陆在林下、空旷地均能生长,说明其在很大的光强范围内可有效地维持光合系统正常运转,但其并没有表现出较高的最大净光合速率,有可能具有较好的光保护机制来应对较强或较弱的光照。基于此,应对美洲商陆的光合作用做深一步的研究,以探明其光适应机理。
致谢:感谢井冈山大学数理学院叶子飘教授在光响应曲线模型方面给予的指导与帮助。
[1] Rober A, Mark A, John S B. Kok effect and the quantum yield of photosynthesis[J]. Plant Physiology, 1984, 75: 95-101.
[2] 叶子飘. 光合作用对光和CO2响应模型的研究进展[J]. 植物生态学报, 2010, 34(6): 727-740.
[3] Baly E C. The kinetics of photosynthesis [J]. Proceedings of the Royal Society of London Series B (Biological Sciences), 1935, 117: 218-239.
[4] Miko UFK, Graham D F. Investigation of the CO2dependence of quantum yield and respiration in eucalyptus pauciflora[J]. Plant Physiology, 1987, 83(4): 1032-1036.
[5] Thornley J H M. Mathematical Models in Plant Physiology[J]. Academic Press, London. 1976, 86-110.
[6] Prioul J L, Chartier P. Partitioning of transfer and carboxylation components of intracellular resistance to photosynthetic CO2fixation: a critical analysis of the methods used[J]. Annals of Botany, 1977, 41:789-800.
[7] Leverenz J W, Jarvus P G. Photosynthesis in Sitka spruce Ⅷ. The effects of light flux density and direction on the rate of net photosynthesis and the stomata conductance of needles[J]. Journal of Applied Ecology, 1979,16: 919-932.
[8] Farquhar G D, Caemmerers S. Berry J A. A biochemical model of photosynthetic CO2assimilation in leaves of C3species[J]. Planta, 1980, 149: 78-90.
[9] Marshall B, Biscoe P V. A model for C3leaves describing the dependence of net photosynthesis on irradiance[J]. Journal of Experiment Botany, 1980, 120:29-39.
[10] Thornley JHM. Dynamic model of leaf photosynthesis with acclimation to light and nitrogen[J]. Annals of Botany, 1998, 81(3): 421-430.
[11] Bassman J, Zwier J C. Gas exchange characteristics of[J]. Tree Physiology, 1991, 8: 145-159.
[12] Prado CHBA, Moraes JAPV. Photosynthetic capacity and specific leaf mass in twenty woody species of Cerrado vegetation under field condition[J]. Photosynthetica, 1997, 33(1): 103-112.
[13] Ye Z P. A new model for relationship between irradiance and the rate of photosynthesis in Oryza sativa[J]. Photosynthetica, 2007, 45(4): 637-640.
[14] Ye Z P, Yu Q, Kang H J. Evaluation of photosynthetic electron flow using simultaneous measurements of gas exchange and chlorophyll fluorescence under photorespiratory conditions[J]. Photosynthetica, 2012, 50(3): 472-476.
[15] White A J, Critchley C. Rapid light curves: A new fluorescence method to asses the state of the photosynthetic apparatus[J]. Photosynthesis Research, 1999, 59: 63-72.
[16] Ralph P J, Gademann R. Rapid light curves: A power fluo tool to assess photosynthetic activity[J]. Aquatic Botany, 2005, 82: 222-237.
[17] 王振兴,朱锦懋,王健,等.闽楠幼树光合特性及生物量分配对光环境的影响[J]. 生态学报, 2012,32(12): 3841-3848.
[18] 孙映波, 操君喜, 尤毅, 等. 不同施肥方式对文心兰生长及光合生理特征的影响[J]. 中国农学通报, 2011, 27(22): 119-124.
[19] 许晨璐, 孙晓梅, 张守攻. 日本落叶松与长白落叶松及其杂种光合特性比较[J]. 北京林业大学学报, 2012, 34(4): 62-66.
[20] 叶子飘, 康华靖. 植物光响应修正模型中系数的生物学意义研究[J]. 扬州大学学报:农业与生命科学版, 2012, 32(2): 51-57.
[21] 户桂敏, 王文天, 彭少麟. 不同氮磷比下入侵种五爪金龙和本地种鸭脚木的竞争表现[J]. 生态环境学报, 2009,18(4): 44-48.
[22] 朱永宁, 张玉书, 纪瑞鹏, 等. 干旱胁迫下3种玉米光响应曲线模型的比较[J]. 沈阳农业大学学报,2012, 43(1): 3-7.
[23] 黄红英, 窦新永, 孙蓓育, 等. 两种不同生态型麻疯树夏季光合特性的比较[J]. 生态学报, 2009,29(6): 2861- 2867.
[24] 王满莲, 冯玉龙, 李新. 紫茎泽兰和飞机草的形态和光合特性对磷营养的响应[J]. 应用生态学报, 2006, 17(4): 602-606.
[25] 廖小锋, 刘济明, 张东凯, 等. 野生小蓬竹的光合光响应曲线及其模型拟合[J]. 中南林业科技大学学报, 2012, 32(3): 124-128.
[26] 侯智勇, 洪伟, 李键, 等. 不同桉树无性系光响应曲线研究[J]. 福建林学院学报, 2009, 29(2): 114-119.
[27] 陈根云, 俞冠路, 陈悦, 等. 光合作用对光合二氧化碳响应的观测方法讨论[J]. 植物生理与分子生物学学报, 2006, 32(6): 691-696.
[28] 刘宇锋, 萧浪涛, 童建华, 等. 非直线双曲线模型在光合光响应曲线数据分析中的应用[J]. 中国农学通报, 2005,121(8): 76-79.
MODEL FITTING ON LIGHT RESPONSE CURVE OF PHOTOSYNTHESIS FOR
*YAN Xiao-hong1.2, HU Wen-hai1, WANG Ning1, ZOU Tan1
(1. School of Life Sciences, Jinggangshan University, Ji’an, Jiangxi 343009, China; 2. College of Forest Resources and Environment Nanjing Forestry University, Nanjing, Jiangsu 210037, China )
In order to investigate applicability of different light-response models to photosynthetic characteristics in, four typical models of light response curve were used to fit the light response curve of its leaf net photosynthesis. The optimal model was screened out by analyzing proximity between the simulated and measured values of photosynthetic parameters. The results showed that light response curves of photosynthesis forall could be fitted by rectangular hyperbolic model, non-rectangular hyperbolic model, index function model and modified rectangular hyperbolic model with determination coefficients of all2>0.99, but only modified rectangular hyperbolic model could calculate their saturation irradiances. According to calculated photosynthetic parameters by various models,n maxestimated by non-rectangular hyperbola was far higher than the measured data, andn maxfitted by exponential function and modified rectangular hyperbolic model were similar with the measured data.satestimated by rectangular hyperbolic model, non-rectangular hyperbolic model and exponential function were far lower than the measured data, onlysatestimated by modified rectangular hyperbolic model were close to the measured data. Therefore, modified rectangular hyperbolic model was the best model in simulating light response curves of photosynthesis and calculate photosynthetic parameters for.n max,sat, andcwhich were calculated by modified rectangular hyperbolic model was respectively 16.98 μmol·m-2·s-1, 1614.54 μmol·m-2·s-1and 14.14 μmol·m-2·s-1, which indicated thatcould be adaptive for illumination intensity in larger range.
L.; light response curve; modified rectangular hyperbolicmodel; saturation irradiance
Q945;S45
A
10.3969/j.issn.1674-8085.2013.03.007
1674-8085(2013)03-0028-06
2013-01-9;
2013-04-06
国家自然科学基金项目(31240015);江西省教育厅科技计划项目(GJJ11717)
*闫小红(1977-),女,内蒙古赤峰人,实验师,南京林业大学博士生,主要从事植物生理生态方面的研究(Email: yanxiaohong325@126.com);
胡文海(1973-),男,江西吉安人,教授,博士,主要从事植物逆境生理方面的研究(Email: huwenhaicy@yahoo.com.cn);
王 宁(1979-),男,山东淄博人,副教授,博士,主要从事植物生态学方面的研究(Email:wangning13@126.com);
邹 谭(1990-),男,江西抚州人,井冈山大学生命科学学院本科生(Email:512974234@qq.com).
猜你喜欢
大众健康(2022年7期)2022-07-30 02:19:04
小哥白尼(野生动物)(2021年12期)2021-03-29 01:15:40
石河子大学学报(自然科学版)(2021年6期)2021-03-03 01:50:22
娃娃乐园·综合智能(2019年10期)2019-12-07 05:27:34
中成药(2018年2期)2018-05-09 07:19:51
新课程·下旬(2017年7期)2017-08-14 11:12:50
首都食品与医药(2015年13期)2015-11-03 08:54:32
小火炬·阅读作文(2014年2期)2015-03-11 04:43:32
江苏中医药(2015年1期)2015-01-09 05:13:31
西北林学院学报(2015年5期)2015-01-03 05:12:20
