
光照强度对许氏平鲉幼鱼的生长、体组分和能量收支的影响*
2013-10-16 07:21房景辉韦秀梅王忠全刘相全宋向军周全利方建光姜海滨
中国海洋大学学报(自然科学版) 2013年9期
房景辉,韦秀梅,王忠全,刘相全,宋向军,周全利,方建光,姜海滨**
(1.山东省海洋水产研究所山东省海洋生态修复重点实验室,山东 烟台264006;2.中国水产科学研究院黄海水产研究所农业部海洋渔业可持续发展重点实验室,山东 青岛266071)
光照是影响生物生长、发育和繁殖的重要因子,对鱼类来说,从仔稚鱼、幼鱼到成鱼等不同的生长发育阶段均受光照影响[1-4]。在光照对鱼类的影响研究中,光照强度对鱼类的生理生态特征影响是一个重要的研究内容[5-6]。鱼类对不同的光照强度经常表现出趋光性和避光性等行为特征,过强和过弱的光线均对鱼类的活动、生长和发育不利[3,7-8]。以前的相关研究主要涉及光照强度对仔稚鱼和早期幼鱼摄食、行为的影响等方面[9-11]。而关于光照强度对鱼类生长、体组分和能量收支的影响研究较少,光照影响鱼类生长的生理生态学机制尚不清楚。
许氏平鲉(Sebastes schlegeli)又称黑鲪,属鲉形目、鲉科、平鲉属,为冷温性近海底层鱼类,卵胎生,洄游范围小,广泛分布于中国的黄、渤和东海。其具有耐温范围广、适应性强、病害少等优点,是利用人工渔礁进行资源增殖和网箱养殖的理想种类[12]。随着人工育苗技术的突破,许氏平鲉逐渐成为中国北方沿海重要的养殖鱼类[13]。目前,关于许氏平鲉的研究已有部分报道,包括营养需求、消化生理、组织生理、摄食生物学和繁殖生物学等[14-17],在基因水平上学者们也做了不少工作[18]。对许氏平鲉生理生态学的研究涉及温度、盐度、运输、体重、密度等环境因子[19-21],但其中很少涉及光照这一重要的环境因子,关于光照强度对其生长和能量分配影响的研究尚未见报道。因此,在许氏平鲉的生产过程中有关光照的操作存在盲目性,生产者选取光照强度往往凭经验,缺乏科学依据,给苗种和成鱼生产造成一定隐患。本研究通过设置不同的光照强度,研究光照强度对许氏平鲉幼鱼的生长、体组分和能量收支的影响,以期查明许氏平鲉幼鱼生长适宜的光照强度范围,从生理和能量学角度揭示光照强度对许氏平鲉幼鱼生长影响的机制。
1 材料与方法
1.1 实验动物来源与驯化
实验用许氏平鲉幼鱼由烟台泰华海珍品有限公司提供,体重(27.35±4.16)g,个体健壮无病,活力强。实验前,将鱼暂养于玻璃钢水槽(1 000L)内,养殖用水为经砂滤的自然海水,盐度31。在20℃下暂养7d。
每天早晚2次投喂配合饵料(山东升索渔用饲料研究中心),其主要生化组成见表1。投喂结束1h后吸底换水,换水率100%/d。光照周期设为14L∶10D,暂养光照强度为700~800lx,24h不间断充气。

表1 饵料的能值及营养组成Table 1 The energy content and composition of the artificial pellet /%
1.2 实验设计与管理
本实验设置7个不同的光照强度处理,即0、100、500、1 000、2 000、4 000和6 000lx,每个梯度下设4个重复。0lx组在隔离的暗室中,仅在实验操作时开灯,每天开灯20~30min,开灯时水槽底部光照强度85lx。其它光照强度组,通过设计不同数量的荧光灯管同时调整灯管与水槽之间的距离获得目标光照强度(以水槽有水时底部中央的光强为准),光照强度以水下照度计(ZDS-10W-2D型)测得。养殖水温由控温仪(WEIPRO MX300 IC)控制,水温波动控制在±0.1℃以内。
暂养结束后,实验鱼饥饿24h使其肠内排空。然后挑选健壮活泼、大小接近的个体,称重后放入相应的水族箱(规格为60cm×40cm×40cm)内进行实验,每个水族箱内各放养4尾幼鱼。同时,每个处理组各取5尾鱼称重后放入-70℃冰箱保存,作为初始样品。
实验期间,每天07:30和18:30分2次投喂配合饲料,根据饵料剩余情况适时调整投喂量。在每次投喂后1h收集残饵和粪便,并于70℃下烘干保存。实际残饵重量通过校正残饵溶蚀率获得。
实验于2011年4月15日开始,到2011年6月14日结束。实验用鱼成活率100%,所有实验鱼饥饿24h后,使其肠内排空,然后进行称量并全部取样用于体组分和能值测定。
1.3 样品测定
实验过程中所取鱼体、粪便以及饲料样品均于70℃下烘干至衡重后称重、研磨,然后进行相关分析。其中,氮含量用凯氏定氮仪(Kjeltec 2100)测定;鱼体和饲料中蛋白质含量由氮含量乘以6.25得到;脂肪测定采用乙醚抽提法;灰分采用马福炉焚烧法(550℃)测定。鱼体、饲料和粪便能值利用PARR6100型氧弹式热量计测定。
1.4 能量收支计算
许氏平鲉幼鱼的摄食(C)、生长(G)、呼吸(R)、粪便(F)和排泄(U)的能量符合下列关系[22]:
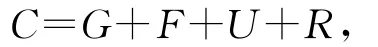
式中:C为摄入的饲料能值;G为生长能;F为排粪能;U为排泄能;R为代谢能;排泄能以下式计算:

式中:CN为摄食食物中所含的氮;GN为鱼体生长增加的氮;FN为粪便中所含的氮;24 830为每克氨氮的能值(J·g-1)[23]。呼吸耗能由能量收支式 R =C-GF-U求出。
1.5 据的计算与处理
实验期间许氏平鲉幼鱼特定生长率、饲料转化率、消化率按如下公式计算:
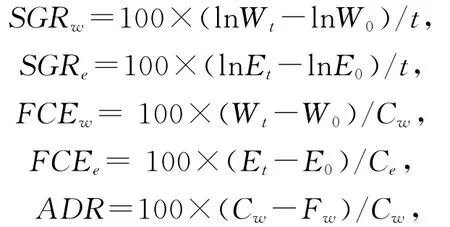
式中:SGRw和SGRe分别为以重量和能量表示的许氏平鲉幼鱼的特定生长率;FCEw和FCEe分别为以重量和能量表示的饲料转化率;ADR为以重量表示的消化率;Cw和Ce分别为以重量和能量表示的许氏平鲉幼鱼的摄食量;Wt和W0为许氏平鲉幼鱼的末体重和初体重;Et和E0为以能量表示的末体重和初体重;t为实验时间。
所得数据用单因素方差分析(ANOVA)及Duncan多重比较进行处理,以P<0.05作为差异显著水平。
2 结果
2.1 生长与摄食
从表2可以看出,各处理组许氏平鲉幼鱼的初始体重没有显著差异(P>0.05),而到实验结束时,光照强度为1 000lx组的幼鱼体重显著高于0、4 000和6 000lx组(P<0.05),在光照强度100~2 000lx间,许氏平鲉幼鱼的末体重没有显著差异(P>0.05)。从日摄食量来看,4 000和6 000lx组的摄食量显著低于500、1 000和2 000lx组(P<0.05)。
各光照处理组许氏平鲉幼鱼的特定生长率见图1,特定生长率随光照强度的增加呈先升高后降低的趋势。各处理组中,1 000lx组的特定生长率最高,该组以体重表示的特定生长率(SGRw)与500和2 000lx组没有显著差异(P>0.05);而以能量表示的特定生长率(SGRe)显著高于其它处理组(P<0.05)。
根据许氏平鲉幼鱼特定生长率随光照强度变化的规律,由回归分析得出其特定生长率(SGRw和SGRe)与光照强度(L)的回归方程:


回归方程中,舍弃了光照强度为0lx组数据。通过回归方程分别得出许氏平鲉幼鱼的最适光照强度为610和630lx。
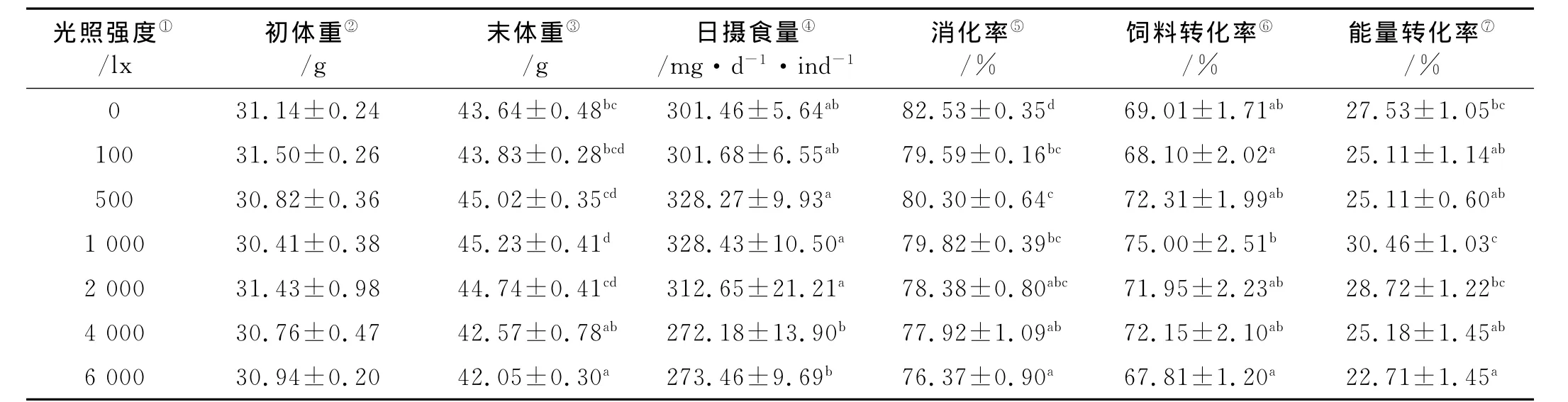
表2 不同光照强度条件下许氏平鲉幼鱼的生长、摄食量、消化率和饲料转化率Table 2 Growth,food consumption,apparent digestion rate,food conversion efficiency and energy conversion efficiency of juvenile Schlegel's rockfish under different light intensity(mean±S.E.)
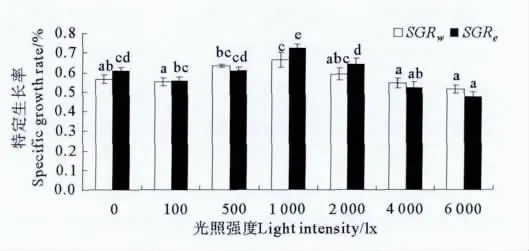
图1 不同光照强度条件下许氏平鲉幼鱼的特定生长率Fig.1 Specific growth rate of juvenile Schlegel’s rockfish under different light intensity during 60dexperiment
2.2 消化率和饲料转化率
由表2可以看出,许氏平鲉幼鱼的消化率随光照强度的增强呈逐渐降低的趋势,以0lx组的消化率最高(P<0.05)。饲料转化率和能量转化率随光照强度的增强均呈先升高后降低的趋势,并且两者的最大值和最小值均分别出现在1 000lx组和6 000lx组。
2.3 体组分和体能值
不同光照强度条件下许氏平鲉幼鱼体组分及体能值见表3。从表3可以看出,许氏平鲉幼鱼体组分中只有灰分含量随光照强度的增强逐渐升高,6 000lx组的灰分含量显著高于0lx组(P<0.05),其它体组分和体能值均不受光照强度的影响,各组之间没有显著差异(P>0.05)。
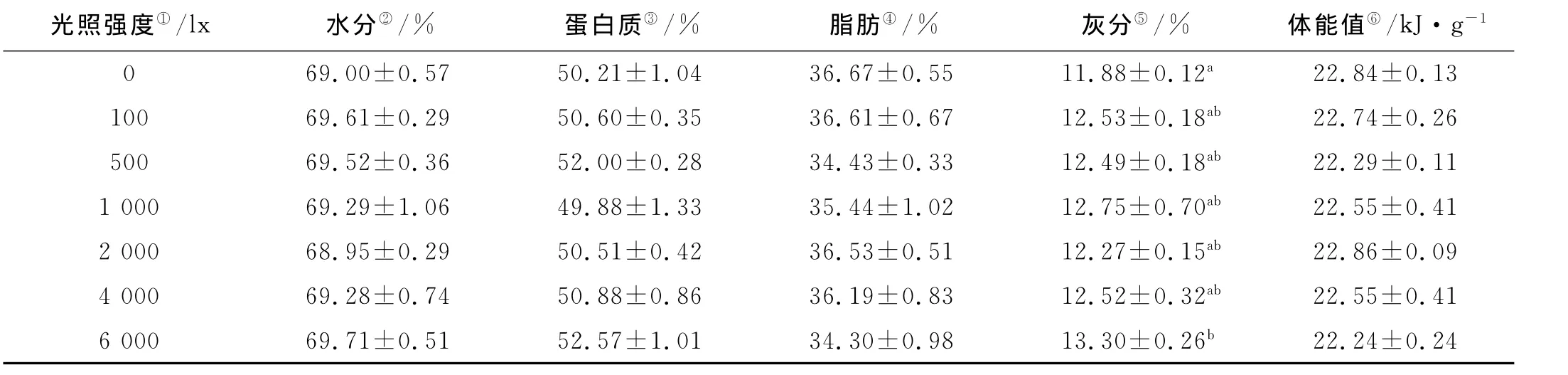
表3 不同光照强度条件下许氏平鲉幼鱼体组分及体能值Table 3 The biochemical composition and body energy content of juvenile Schlegel’s rockfish under different light intensity
2.4 能量收支
从表4可以看出,不同光照强度条件下许氏平鲉幼鱼能量收支各组分能值均随光照强度的增强呈先升高后降低的趋势。1 000lx组的摄食能、生长能和排泄能均高于其它处理组,但是其摄食能与500和2 000lx组没有显著差异(P>0.05),其排泄能与1 000lx以下光照处理组没有显著差异(P>0.05),而1 000lx组的生长能显著高于其它处理组(P<0.05)。500lx组的粪能和呼吸能在各组中最高。
从能量收支情况(见表5)看,光照强度为1 000lx组的幼鱼生长能占摄食能的比例为31.99%,显著高于其它处理组,6 000lx组幼鱼的生长能占摄食能的比例最低。与生长能相对应,1 000lx组幼鱼的呼吸能占摄食能的比例最低,而6 000lx组幼鱼的呼吸能占摄食能的比例最高,为61.76%。各处理中,粪能占摄食能的比例随光照强度变化趋势不明显,0lx组幼鱼的粪能占摄食能的比例最低(P<0.05)。排泄能占摄食能的比例受光照强度影响不显著(P>0.05)。

表4 不同光照强度条件下许氏平鲉幼鱼能量收支各组分能值Table 4 Energy parameters of juvenile Schlegel’s rockfish under different light intensity during 60dexperiment/J·g-1·d-1

表5 不同光照强度条件下许氏平鲉幼鱼的能量收支Table 5 Energy allocation of juvenile Schlegel’s rockfish under different light intensity during 60dexperiment /%
3 讨论与分析
3.1 许氏平鲉幼鱼生长的最适光照强度
光照强度作为一个重要的环境因子,可以对鱼类的摄食和生长造成显著影响[9-10,24]。李修峰等[10]发现大眼鳜(Siniperca kneri)幼鱼(32~80g)喜欢在黑暗环境中摄食,其在黑暗环境中的摄食量分别是自然光和强光环境中的1.1和2.0倍。花鲈(Lateolabrax j aponicus)幼鱼((3.4±0.2)cm)在光照强度为400lx的环境中摄食强度显著高于在其它光强梯度下的摄食强度[24]。而在103~10-3lx的光照强度范围内,乌鳢(Channa argus)幼鱼(26.1~32.0mm)对浮游动物的摄食强度随光照强度的减弱而增强,并在10-3lx达到最 大 值[25]。 褐 牙 鲆 (Paralichthys olivaceus)幼 鱼((2.21±0.22)cm)在15d的光照强度实验中,500lx光照强度组摄食最佳,并且获得了29.0%的体长增长率,摄食和生长均显著高于其它光照强度处理组(P<0.05)[9]。在本研究中,在光照强度低于2 000lx处理组中许氏平鲉幼鱼的日摄食量没有显著差异(P>0.05),而4 000和6 000lx组幼鱼的日摄食量显著低于500、1 000和2 000lx组(P<0.05)。从摄食情况来看,许氏平鲉幼鱼对光照强度具有较强的适应能力,而强光照(4 000lx以上)仍然是其摄食的不利因素。这一研究结果与之前的报道有所差异,这些差异表明不同种类对光照强度的适应性差异较大,而这些研究中涉及鱼类的生长阶段不同也可能是差异产生的重要原因。与摄食情况相似,本研究中100~2 000lx各组中幼鱼的末体重没有显著差异,说明许氏平鲉幼鱼生长适宜的光照强度变化幅度较大,这与以前研究中褐牙鲆幼鱼生长的适宜光照强度较窄的研究结果不同[9],而与乌鳢幼鱼对光照的适应范围较广相似[25]。而通过不同光照强度下许氏平鲉幼鱼(30~45g)的特定生长率与光照强度的回归方程,可以得出其生长最适的光照强度为610~630lx,由此可以确定许氏平鲉幼鱼喜弱光的生活习性。因此,鱼类虽然对光照强度有一定的适应能力,但是较弱的光照强度仍然是多数种类在幼鱼时期的适宜生活环境之一。鱼类的这种习性可能与其在水中的自然生活环境相对应,是否如此,需要进一步研究证实。
3.2 光照强度对许氏平鲉幼鱼体组分的影响
国内外关于光照强度对鱼类体组分和体能值影响的研究报道较少[26]。而以往的研究表明,光照可以影响甲壳动物的体组分[26-27]。Li等发现中华绒螯蟹(Eriocheir sinensis)的体组分受光照强度影响显著,脂肪含量随光照强度的减弱而降低,不同光照强度处理组的蛋白和灰分含量均差异显著(P<0.05),其推测体组分的差异是由体内某些酶活力不同而导致[26]。在本研究中,体组分和体能值受光照强度的影响不显著,这一现象在以往有关环境因子对鱼类体组分的研究中少见报道[28-29],这说明光照强度是一个比较缓和的环境因子,许氏平鲉幼鱼对光照强度具有较广的适应范围和较强的耐受能力。至于许氏平鲉幼鱼如何通过生理活动来维持体组分和体能值的恒定,尚需进一步研究。
3.3 光照强度影响许氏平鲉幼鱼生长的生物能量学机制
很多研究证明光照对鱼类的生理活动可以产生多方面影响[30],光照强度可以影响虹鳟(Salmo gairdneri)血浆中甲状腺素的含量[5]。本研究中光照强度对许氏平鲉幼鱼的消化率、饲料转化率和能量转化率均产生显著影响,生长最佳的1 000lx组具有较高的消化率和转化率,反之,生长最差的6 000lx组其消化率和转化率也最低。在以往对许氏平鲉能量收支的研究中[20,31],呼吸能和生长能主导了能量分配模式,这与本研究结果类似,在本研究中1 000lx光照组生长能占摄食能的比例最高,而6 000lx组的呼吸能占摄食能的比例最高,这说明高光照强度条件下,许氏平鲉幼鱼用于代谢的能量较多,从而用于生长的能量减少。这一点与大多数甲壳动物类似,在高光照条件下具有较高的代谢[32]。因此,不同光照强度条件下,许氏平鲉幼鱼的消化率和转化率不同是导致其生长差异的重要原因,而高光照强度条件下其用于呼吸代谢的能量增多是其生长减缓的机制之一。
4 结语
综上所述,在本实验条件下,许氏平鲉幼鱼生长最适的光照强度范围为610~630lx,说明其具有喜弱光的生活习性,但在100~2 000lx的光强范围内其生长不受影响。虽然光照强度可显著影响许氏平鲉幼鱼的生长、摄食、消化率和饵料转化率等,但是其体组分和体能值不受光照强度的影响,这说明在本研究所涉及的光照强度范围内,许氏平鲉幼鱼的主要营养成分不受光照强度的影响,其对光照强度具有较强的适应能力。据此,在养殖生产中,可以通过适当遮挡光线或调整网箱水深获得许氏平鲉幼鱼生长的适宜光照强度。同时,搭建人工鱼礁可以为许氏平鲉幼鱼创造更多适宜栖息的光照环境。
[1] Yadav M,Ooi S D.Effect of photoperiod on the incorporation of 3H-thymidine into the gonads of Carassius auratus[J].Journal of Fish Biology,1977,11(5):409-416.
[2] Davies P R,Hanyu I,Furukawa K,et al.Effect of temperature and photoperiod on sexual maturation and spawning of the common carpⅢ.Induction of spawning by manipulating photoperiod and temperature[J].Aquaculture,1986,52(2):137-144.
[3] Cuvier-Péres A,Jourdan S,Fontaine P,et al.Effects of light intensity on animal husbandry and digestive enzyme activities in sea bass Dicentrachus labrax post-larvae[J].Aquaculture,2001,202(3-4):317-328.
[4] 孙中之,柳学周,徐永江,等.半滑舌鳎工厂化人工育苗工艺技术研究[J].中国水产科学,2007,14(2):244-248.
[5] Cook R F,Eales J G.Effects of feeding and photocycle on diel change in plasma thyroid hormone levels in rainbow trout,Salmo gairdneri[J].Journal of Experimental Zoology,1987,242(2):161-169.
[6] 王武,李伟纯,马旭洲,等.水温与光照对瓦氏黄颡鱼幼鱼行为的影响[J].生态学杂志,2008,27(5):791-796.
[7] 何大仁,蔡厚才.鱼类行为学[M].厦门:厦门大学出版社,1998.
[8] Browman H I,Skiftesvik A B,Kuhn P.The relationship between ultraviolet and polarized light and growth rate in the early larval stages of turbot (Scophtalmus maximus),Atlantic cod (Gadus morhua)and Atlantic herring(Clupea harengus)reared in intensive culture conditions[J].Aquaculture,2006,256(1-4):296-301.
[9] 楼宝,史会来,毛国民,等.不同光照时间、强度对舟山褐牙鲆稚幼鱼摄食、生长的影响[J].浙江海洋学院学报,2008,27(2):140-143.
[10] 李修峰,黄道明,杨汉运.光照对大眼鳜鱼幼鱼摄食强度的影响[J].湖南农业大学学报,2005,31(2):187-190.
[11] Almazán-Rueda P,Schrama J W,Verretha J A J.Behavioural responses under different feeding methods and light regimes of the African catfish (Clarias gariepinus)juveniles[J].Aquaculture,2004,231(1-4):347-359.
[12] 陈大刚,叶振江,段钰,等.许氏平鲉繁殖群体的生物学及其苗种培育的初步研究[J].海洋学报,1994,16(3):94-101.
[13] 姜海滨,杜荣斌,鹿叔锌,等.温度、盐度对许氏平鲉幼鱼存活及生长的影响[J].齐鲁渔业,2010,27(6):9-10.
[14] Nakagawa M,Hirose K.Individually specific seasonal cycles of embryonic development in cultured broodstock females of the black rockfish,Sebastes schlegeli[J].Aquaculture,2004,233:549-559.
[15] Lim S R,Choi S M,Wang X J,et al.Effects of dehulled soybean meal as a fish meal replacer in diets for fingerling and growing Korean rockfish Sebastes schlegeli[J].Aquaculture,2004,231:457-468.
[16] 万军利.野生与养殖黑鲪消化酶活力的比较[J].生态学杂志,2010,29(5):1035-1038.
[17] 严全根,解绶启,雷武,等.许氏平鲉幼鱼的赖氨酸需求量[J].水生生物学报,2006,30(4):459-465.
[18] Kim D,Cho Y C,Sohn Y C.Molecular characterization of rock-fish(Sebastes schlegeli)gonadotropin subunits and their mRNA expression profiles during oogenesis[J].General and Comparative Endocrinology,2005,141(3):282-290.
[19] Jeon J,Lim H K,Kannan K,et al.Effect of perfluorooctanesulfonate on osmoregulation in marine fish,Sebastes schlegeli,under different salinities[J].Chemosphere,2010,81(2):228-234.
[20] 孙耀,张波,郭学武,等.体重对黑鲪能量收支的影响[J].海洋水产研究,1999,20(2):66-70.
[21] 洪磊.环境胁迫对鱼类生理机能影响的初步研究[D].青岛:中国海洋大学,2004.
[22] Carfoot T H.Animal Energetics[M].New York:New York Academic Press,1987:407-515.
[23] 崔奕波.鱼类生物能量学的理论与方法 [J].水生生物学报,1989,13(4):369-383.
[24] 姜志强,谭淑荣.不同光照强度对花鲈幼鱼摄食的影响[J].水产科学,2002,21(3):4-5.
[25] 谢从新,熊邦喜,周洁,等.不同照度下乌鳢幼鱼摄食强度及其动力学[J].水生生物学报,1997,21(3):213-217.
[26] Li X,Li Z,Liu J,et al.Effects of light intensity on molting,growth,precocity,digestive enzyme activity,and chemical composition of juvenile Chinese mitten crab Eriocheir sinensis[J].Aquaculture International,2011,19:301-311.
[27] Teschke M,Kawaguchi S,Meyer B.Effects of simulated light regimes on maturity and body composition of Antarctic krill,Euphausia superba[J].Marine Biology,2008,154:315-324.
[28] 王晓杰,张秀梅,黄国强.低温胁迫对许氏平鲉补偿生长的影响[J].中国水产科学,2006,13(4):566-572.
[29] 房景辉,田相利,董双林,等.温度对半滑舌鳎(Cynoglossus semilaevis)的生长、生化组成和能量收支的影响[J].中国海洋大学学报:自然科学版,2010,40(1):25-30.
[30] 周显青,牛翠娟,李庆芬.光照对鱼类生理活动影响的研究进展[J].生态学杂志,1999,18(6):59-61.
[31] 孙耀,郑冰,张波,等.温度对黑鮶(Sebastodes fuscescens)能量收支的影响[J].海洋学报,2003,25(2):196-201.
[32] Hoang T,Barchiesis M,Lee S Y,et al.Influences of light intensity and photoperiod on moulting and growth of Penaeus merguiensis under laboratory condition[J].Aquaculture,2003,216,(1-4):343-354.
猜你喜欢
林业与生态科学(2022年4期)2022-12-24
轻工学报(2022年5期)2022-10-08
上海蔬菜(2022年2期)2022-04-29
中国海洋大学学报(自然科学版)(2021年1期)2021-12-02
水产养殖(2021年5期)2021-05-21
防护林科技(2020年9期)2020-11-09
养生保健指南(2018年4期)2018-04-11
中南林业调查规划(2017年3期)2017-12-29
养生保健指南(2016年12期)2017-01-06
现代农业科技(2009年19期)2009-03-20
