
猪鼻支原体vlp重复区融合蛋白的构建表达及反应原性分析
2013-09-26 02:55熊祺琰刘占军马庆红冯志新刘茂军白方方邵国青
中国人兽共患病学报 2013年9期
熊祺琰,纪 燕,刘占军,马庆红,冯志新,刘茂军,白方方,邵国青
猪鼻支原体(Mycoplasma hyorhinis)(以下简写为Mhr)是一种能够引起猪多发性浆膜炎、关节炎、中耳炎、肺炎等病症的致病性支原体[1-4]。Mhr在临床猪场的感染率较高,通常由母猪或大猪传染给小猪,主要通过上呼吸道感染传播,该病原从呼吸系统向全身发生侵染将导致多浆膜炎、关节炎等疾病的发生。Mhr也是细胞培养中引起培养污染的最常见支原体[5]。近年来的研究发现Mhr感染与人的多种癌症,包括胃癌、大肠癌、前列腺癌、胰腺癌、肺癌等有明显的相关性[6-9]。目前临床上 Mhr的感染检测主要依赖于PCR方法,尚未有商品化试剂盒用于血清学检测。
支原体的基因组相对较小,缺乏细胞壁结构,在多种支原体的基因组中都含有多个编码表面可变脂蛋白的基因,其功能主要通过改变脂蛋白种类及大小来改变支原体表面的抗原性,帮助支原体逃避宿主的免疫系统识别,从而在不同宿主中实现持续感染。Mhr中发现的表面可变脂蛋白为vlp(variable lipoprotein,vlp)[10-11],目前发现的 Mhr vlp家族成员共有7个,分别为vlpA-vlpG。本研究拟构建表达一种重组蛋白,包含vlpA-vlpG的主要表位,该重组蛋白可用于建立Mhr的血清学诊断方法,及用于开发Mhr重组蛋白疫苗。
1 材料与方法
1.1 试验材料
1.1.1 菌种与质粒 大肠杆菌 BL-21(DE3)及表达质粒载体pET-32a(+)均由江苏省农业科学院兽医研究所保存。
1.1.2 材料与试剂 硝酸纤维素膜,美国 Millipore公司;镍柱,金斯瑞生物科技公司;辣根过氧化物酶标记的羊抗猪IgG,美国Bethyl公司;辣根过氧化物酶标记的羊抗兔IgG,武汉博士德生物工程有限公司;DAB显色液,武汉博士德公司;基因合成,金斯瑞生物科技公司。Mhr阳性兔血清,实验室自制,用Mhr全菌灭活抗原免疫5只健康新西兰大白兔,免疫3次每次间隔2周,第3次免疫后2周采血;Mhr阳性猪血清,实验室自制,用Mhr全菌灭活抗原免疫3头健康猪,免疫3次每次间隔2周,第3次免疫后2周采血。
1.2 试验方法
1.2.1 vlpA-G蛋白设计 在GenBank上检索vlp家族成员序列,用blast分析蛋白片段的Mhr特异性,并分析各自的重复区序列。为维持表位的完整性,vlpB-vlpG蛋白的重复区都采用2段重复片段。各个序列之间添加柔性片段GGGGS作为连接肽。由于vlpA可能仅含有两段重复区片段并间隔较远,故仅取一段vlpA的重复片段,以GGGGS为连接肽串联在vlpB-G的C端,设计得到重组蛋白vlpA-G。
1.2.2 重组蛋白vlpA-G表达质粒的构建 采用基因合成的方式获得表达vlpB-G的基因序列,同时利用大肠杆菌偏爱的密码子进行序列优化以帮助表达。两端分别添加EcoRⅠ和HindⅢ酶切位点。合成的基因片段经EcoRⅠ和HindⅢ双酶切后插入pET-32a(+)空质粒载体中,构建得到 pET32avlpB-G重组表达质粒。
在vlpB-G序列的基础上,根据GenBank中登录的vlpA序列(登录号为AF193874.1)设计引物扩增vlpA基因重复片段放入vlpB-G的C端,其间加入GGGGS作为连接肽。具体方法为:以vlpB-G基因为模板用下游两次加端PCR的方法扩增得到vlpA-G基因,C端加入HindⅢ酶切位点。PCR引物分别为:上游引物,5′-AAAGAATTCGGCGGTGGCGGCTCGGGCACGGGCTCGGATTCTCAG 3′;下游引物-1,5′-TGCGTATTTTCGGTTTTCG AGCCGCCACCGCCGCTGTCTGCCTGACCGCT GCTTT 3′;下游引物-2,5′-CCGAAGCTTAGGTGCCCGGCGCCTCGCTTTGCTGCGTATTTTCG GTTTTCGAGCC 3′。扩增的PCR产物经EcoRⅠ和HindⅢ双酶切后插入pET-32a(+)空质粒载体中,构建得到表达vlpA-G的重组质粒pET32a-vlpA-G。转化大肠杆菌BL21感受态细胞,用上游引物和下游引物-2PCR筛选阳性克隆,得到的阳性克隆进行一次单克隆,再重复筛选一次,获得阳性菌株,测序鉴定序列的正确性。制备甘油管保存菌种。
1.2.3 重组vlpA-G蛋白的表达 接种20μL甘油管菌液至5mL氨苄抗性的LB培养基中,过夜培养得到种子液,次日将种子液以2%的量转接至氨苄抗性的LB培养基中,振荡培养至OD600值为0.4~0.6,加IPTG至终浓度1mmol/L,37℃诱导,于不同时间点收集菌体,进行SDS-PAGE(15%分离胶、5%浓缩胶),测定最佳诱导时间。
1.2.4 重组vlpA-G蛋白的制备及纯化 同上接种,并于上述摸索的最佳诱导时间点收集菌体,超声裂解菌体,离心收集上清,镍柱亲和层析纯化。
1.2.5 Western Blot检测 Mhr抗体 蛋白经SDS-PAGE分离,用半干转印法(Tris-Gly缓冲系统,0.65mA/cm2,转印1.5h)转移至硝酸纤维素膜(NC)上。5%脱脂奶室温或37℃摇动封闭2h,加入用1% 脱脂奶粉,10mmol/L PBS稀释40倍的Mhr阳性兔血清,37℃摇动杂交1h。0.05%Tween-20,10mmol/L PBS洗膜5次,每次5min。洗涤后加入用1% 脱脂奶粉,10mmol/L PBS稀释5 000倍的羊抗兔酶标二抗,37℃作用1h。同上洗涤后加入显色液,出现明显条带后用蒸馏水冲洗膜以终止反应。
1.2.6 间接ELISA方法建立 取纯化的重组vlpA-G蛋白,考马斯亮蓝试剂盒测定蛋白浓度。用包被稀释液0.05mmol/L Na2CO3-NaHCO3缓冲液(pH 9.6)将蛋白稀释至为5μg/mL,每孔加100 μL,4℃包被过夜,次日用含0.05%Tween-20的PBST溶液洗涤3次,每次5min;每孔加200μL 3%BSA于37℃封闭2h,同上洗涤3次;加入1∶100倍稀释的Mhr抗体阳性猪血清或阴性血清样本,每孔100μL,置37℃孵育1h,洗涤;加入1∶8 000稀释的辣根过氧化物酶标记的羊抗猪IgG二抗,每孔100μL,置37℃孵育1h,洗涤;加新鲜配制的TMB底物100μL,37℃避光显色10min;加入50μL 2 mol/L H2SO4终止反应,于450nm波长测定。
2 结 果
2.1 vlpA-G蛋白设计 在GenBank上检索vlp家族成员序列,7种vlp蛋白vlpA、vlpB、vlpC、vlpD、vlpE、vlpF、vlpG结构及重复片段区代表性示意图如图1。
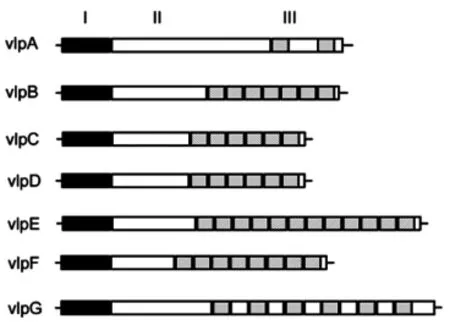
图1 vlp家族蛋白Fig.1 The vlp family protein
经Blast分析发现vlp蛋白都具有较强的Mhr特异性,适合作为检测抗原使用。将vlpB、vlpC、vlpD、vlpE、vlpF、vlpG各自的2段重复片段串联,之间添加柔性片段GGGGS作为连接肽,设计重组蛋白vlpB-G(图2A)。取vlpA的一段重复片段串联至vlpB-G的C端,中间添加柔性片段GGGGS作为连接肽,设计得到重组蛋白vlpA-G。具体蛋白序列如图2B所示。
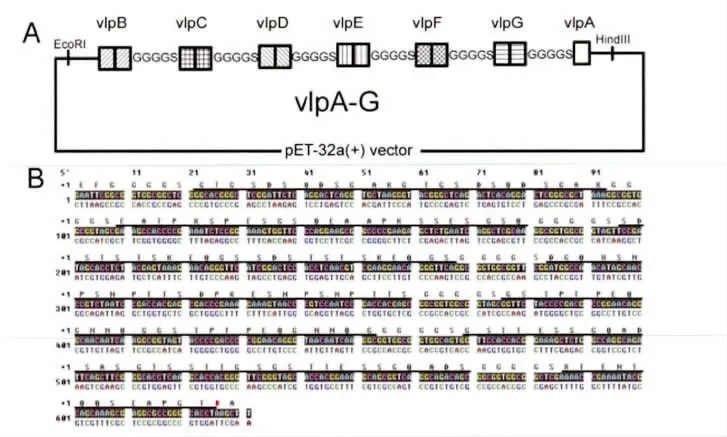
图2 重组vlpA-G蛋白设计及其序列Fig.2 Design and sequence of the recombinant vlpA-G protein
2.2 重组表达质粒pET32a-vlpA-G的构建 采用基因合成的方式获得串联表达vlpB-vlpG重复区多肽的基因序列,用加端PCR方法将vlpA重复片段基因串联至vlpB-G基因C端,插入pET32a-vlpB-G重组质粒中,构建得到表达vlpA-G的重组质粒pET32a-vlpA-G。用vlpA-G上下游引物进行PCR筛选(图3),挑选阳性克隆,经单克隆后DNA测序,测序结果正确。制备甘油管保存菌种。
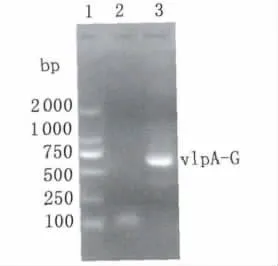
图3 重组表达质粒pET32a-vlpA-G的鉴定1.DNA分子量标准;2.阴性对照;3.vlpA-G基因扩增产物Fig.3 Identification of the recombinant plasmid expressing vlpA-G1:DNA molecular weight markers;2:Negative control;3:PCR product of vlpA-G gene
2.3 重组蛋白vlpA-G的表达及分离纯化 IPTG诱导后重组vlpA-G蛋白可以在大肠杆菌中有效地表达,在分子量为50kDa与90kDa的标准条带位置中间偏50kDa处有显著加深的蛋白带。IPTG诱导后不同时间取样,发现加入IPTG诱导6h后蛋白的表达达到高峰并处于稳定水平,故选择诱导6h后收集菌体。采用镍柱亲和纯化重组vlpA-G蛋白,结果见图4。对蒸馏水透析后冻干保存于-20℃。
2.4 Western Blot检测 Mhr抗体 将纯化的vlpA-G重组蛋白以及pET-32a(+)空载体表达纯化蛋白电泳分离后用Mhr抗血清进行 Western Blot检测,结果可见vlpA-G重组蛋白可与Mhr阳性兔血清产生明显杂交条带,而pET-32a(+)空载体表达纯化蛋白与Mhr阳性兔血清无杂交反应,说明重组vlpA-G蛋白具有明显的Mhr抗体反应原性。以未免疫健康兔血清为阴性对照,未发现杂交条带。
2.6 间接ELISA检测Mhr抗体 以重组蛋白vlpA-G为包被抗原检测Mhr免疫猪血清,以未免疫健康猪血清为阴性对照。结果表明Mhr免疫猪血清中可检测到明显的Mhr抗体,与阴性血清相比差异极显著,说明该重组vlpA-G蛋白具有良好的反应原性,可作为候选抗原蛋白用于血清学诊断方法的后续研究。

图4 重组vlpA-G蛋白的表达纯化1.蛋白分子量标准;2.pET32a-vlpA-G 重组菌诱导后;3.pET-32a(+)空载体对照菌诱导后;4.纯化的vlpA-GFig.4 Expression and purification of the recombinant vlpA-G protein1:Protein molecular weight markers;2:Total protein of E.coli BL-21containing pET32a-vlpA-G after induction;3:Total protein of E.coli BL-21containing pET-32a(+)after induction;4:Purified vlpAG protein
3 讨 论
Mhr是第一种从猪体内分离到的支原体,主要通过呼吸道感染传播,但目前Mhr的研究较少,其具体的致病机制尚不清楚,可能通过黏附到纤毛细胞上进而侵入机体[12-13],同时 Mhr对纤毛的损伤也为其他病原菌的感染提供便利从而导致共感染的发生。除了引起天然宿主猪的疾病外,Mhr也被发现与多种人类肿瘤相关。早在上世纪90年代人们就注意到某些支原体的感染与人类肿瘤之间存在着一定的相关性,其中包括 Mhr。Mhr的一种膜蛋白P37被证明与细胞肿瘤化有直接关系[8,14],是目前唯一鉴定出的Mhr肿瘤相关抗原。
目前,猪群Mhr感染的诊断手段主要包括分离培养、PCR[15]、免 疫 组 化[16]及 原 位 杂 交[3]。 利 用PCR检测16sRNA、P37基因建立PCR方法能够较为灵敏地检测Mhr,并能与猪滑液支原体、猪肺炎支原体区别开来。但活体动物的PCR通常使用鼻拭子及支气管肺泡灌洗液作为待检样本。其中鼻拭子样本检出率较低,通常只能用于群体感染性的判断,而对于个体发病而言容易出现假阴性。支气管肺泡灌洗液虽然可以通过麻醉动物后利用纤维支气管镜等仪器实现活体采集,但是难以应用于临床进行大规模采样。免疫组化及原位杂交方法虽然也可以特异性地检测猪体内的Mhr,但是同样都需要剖杀动物检查。血清学检测是临床主要依赖的检测手段。刘星等报道利用Mhr特异性的单抗建立阻断ELISA方法可用于检测血清中Mhr[17]。
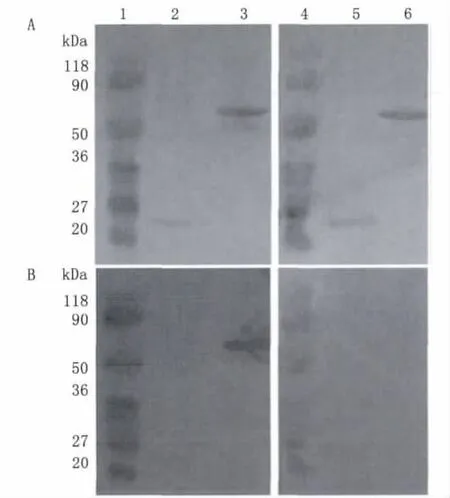
图5 重组蛋白vlpA-G Western Blot检测 Mhr血清抗体A:丽春红染色结果;B:Western blot杂交结果1-3.Mhr阳性血清杂交;4-6.Mhr阴性血清杂交1、4泳道,蛋白分子量标准;2、5泳道,转化pET-32a(+)空载体的工程菌纯化表达蛋白;3、6泳道,纯化重组蛋白vlpA-G.Fig.5 Detection of serum antibody against Mhr by recombinant vlpA-G protein in Western blot assayA:Stained by poncean before hybridization;B:Result of Western blot1-3:Hybridized with Mhr positive serum;4-6:Hybridized with Mhr negative serum;1and 4:Protein molecular weight markers;2and 5:Purified protein of E.coli BL-21containing pET-32a(+)after induction;3and 6:Purified recombinant vlpA-G protein.
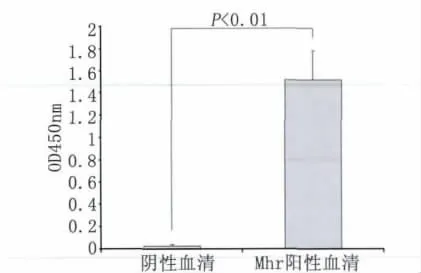
图6 重组蛋白vlpA-G ELISA检测Mhr血清抗体Fig.6 Detection of serum antibody against Mhr by the ELISA assay using recombinant vlpA-G protein as coating antigen
本研究利用Mhr vlp可变脂蛋白家族为靶蛋白,构建重组蛋白拟作为ELISA包被抗原使用。Mhr vlp家族蛋白结构类似都由3部分构成[10-11]:I区为信号肽区域,7种vlp蛋白的信号肽区序列完全相同;II区为相对保守的过渡区,III区为重复片段区,由12~13个氨基酸的短肽片段串联重复而成,重复次数2~11个不等,7种vlp蛋白重复序列各不相同,少数vlp蛋白在重复片段之间存在连接序列。在不同菌株及同一菌株传代过程中vlp蛋白家族表达的数量、种类和分子量大小各不相同,这种表达差异使得Mhr具有多变的表面抗原性,在免疫逃避中起到关键作用。vlp家族蛋白作为Mhr的表面脂蛋白,是建立血清学诊断方法的理想抗原,同时表面蛋白通常也具有较好的免疫原性,因此也适合作为重组疫苗的候选蛋白。介于vlp家族蛋白本身的表达变异性,在本研究中设计的vlpA-G串联表达了vlp家族7种蛋白的重复片段,囊括了目前已知的vlp家族的所有成员的主要免疫区段,可以对广泛的Mhr菌株产生反应。在试验过程中我们发现表达出来的vlpA-G蛋白的分子量大于50kDa,而设计的重组蛋白理论分子量应为36.8kDa,另一方面,DNA测序结果却显示所设计的vlpA-G基因序列正确地插入到pET-32a(+)载体中。其分子量与预期不符的原因可能与vlp蛋白本身的功能和性质有关,在其他支原体的一些黏附蛋白上也报道过这种现象[18]。
本研究结果证明,重组vlpA-G可以在大肠杆菌中很好地表达,以其为包被抗原初步建立ELISA方法能够明显地检测到Mhr的阳性血清。本结果将为Mhr血清或黏膜抗体间接ELISA检测方法的建立奠定基础。
[1]Lin JH,Chen SP,Yeh KS,etal.Mycoplasma hyorhinis in Taiwan:diagnosis and isolation of swine pneumonia pathogen[J].Vet Microbiol,2006,115(1-3):111-116.DOI:10.1016/j.vetmic.2006.02.004
[2]Morita T,Fukuda H,Awakura T,etal.Demonstration of Mycoplasma hyorhinis as a possible primary pathogen for porcine otitis media[J].Vet Pathol,1995,32(2):107-111.DOI:10.1177/030098589503200202
[3]Kim B,Lee K,Han K,etal.Development of in situ hybridiza-tion for the detection of Mycoplasma hyorhinis informalin-fixed paraffin-embedded tissues from naturally infected pigs with polyserositis[J].J Vet Med Sci,2010,72(9):1225-1227.DOI:10.1292/jvms.10-0062
[4]Jayagopala RN,Wilkie BN,Borgs P,etal.Cytokines in Mycoplasma hyorhinis-induced arthritis in pigs bred selectively for high and low immune responses[J].Infect Immun,2000,68(3):1150-1155.DOI:10.1128/IAI.68.3.1150-1155.2000
[5]Zinocker S,Wang MY,Gaustad P,etal.Mycoplasmacontamination revisited:mesenchymal stromal cells harboring Mycoplasma hyorhinis potently inhibit lymphocyte proliferation in vitro[J].PLoS One,2011,6(1):e16005.DOI:10.1371/journal.pone.0016005
[6]Yang H,Qu L,Ma H,etal.Mycoplasma hyorhinis infection in gastric carcinoma and its effects on the malignant phenotypes of gastric cancer cells[J].BMC Gastroenterol,2010,10:132.DOI:10.1186/1471-230X-10-132
[7]Namiki K,Goodison S,Porvasnik S,etal.Persistent exposure to Mycoplasmainduces malignant transformation of human prostate cells[J].PLoS One,2009,4(9):e6872.DOI:10.1371/journal.pone.0006872
[8]Urbanek C,Goodison S,Chang M,etal.Detection of antibodies directed at M.hyorhinis p37in the serum of men with newly diagnosed prostate cancer[J].BMC Cancer,2011,11:233.DOI:10.1186/1471-2407-11-233
[9]Mariotti E,Gemei M,Mirabelli P,etal.The percentage of CD133+ cells in human colorectal cancer cell lines is influenced by Mycoplasma hyorhinis infection[J].BMC Cancer,2010,10:120.DOI:10.1186/1471-2407-10-120
[10]Citti C,Watson-McKown R,Droesse M,etal.Gene families encoding phase-and size-variable surface lipoproteins of Mycoplasma hyorhinis[J].J Bacteriol,2000,182(5):1356-1363.DOI:10.1128/JB.182.5.1356-1363.2000
[11]Citti C,Kim MF,Wise KS.Elongated versions of Vlp surface lipoproteins protect Mycoplasma hyorhinis escape variants from growth-inhibiting host antibodies[J].Infect Immun,1997,65 51773-1785.
[12]Johannsen U,Menger S,Kinne J,etal.The pathology and pathogenesis of experimental Mycoplasma hyorhinis infection of piglets with and without thermomotor stress.2.Electron microscopic studyresults[J].Zentralbl Veterinarmed A,1991,38(5):321-336.
[13]Morita T,Fukuda H,Awakura T,etal.Demonstration of Mycoplasma hyorhinis as a possible primary pathogen for porcine otitis media[J].Vet Pathol,1995,32(2):107-111.DOI:10.1177/030098589503200202
[14]Gong M,Meng L,Jiang B,etal.p37from Mycoplasma hyorhinis promotes cancer cell invasiveness and metastasis through activation of MMP-2and followed by phosphorylation of EGFR[J].Mol Cancer Ther,2008,7(3):530-537.DOI:10.1158/1535-7163.MCT-07-2191
[15]Caron J,Ouardani M,Dea S.Diagnosis and differentiation of Mycoplasma hyopneumoniae and Mycoplasma hyorhinis infections in pigs by PCR amplification of the p36and p46genes[J].J Clin Microbiol,2000,38(4):1390-1396.
[16]Morita T,Muraki Y,Awakura T,etal.Detection of Mycoplasma hyorhinis in porcine eustachitis[J].J Vet Med Sci,1993,55(3):475-477.DOI:10.1292/jvms.55.475
[17]Liu X,Zhang LF,Yang YP,etal.Preparation of monoclonal antibodies of Mycoplasma hyorhinis and establishment of antibody-sandwich ELISA method[J].Chin J Comp Med,2008,18(3):55-58.(in Chinese)刘星,张丽芳,杨玉萍,等.抗猪鼻支原体单克隆抗体的研制及双抗体夹心ELISA检测法的建立[J].中国人兽共患病学报,2008,18(3):55-58.DOI:10.3969/j.issn.1671-7856.2008.03.013
[18]Jenkins C,Wilton JL,Minion FC,etal.Two domains within the Mycoplasma hyopneumoniae cilium adhesin bind heparin[J].Infect Immun,2006,74(1):481-487.DOI:10.1128/IAI.74.1.481-487.2006
猜你喜欢
广东药科大学学报(2022年3期)2023-01-04
生物学通报(2022年1期)2022-11-22
成都医学院学报(2022年4期)2022-08-19
保健与生活(2022年16期)2022-08-06
江西农业学报(2021年4期)2021-04-20
中华养生保健(2020年2期)2020-11-16
三农资讯半月报(2020年11期)2020-06-21
中国现代医药杂志(2020年12期)2020-02-06
生物学教学(2019年3期)2019-03-22
中国当代医药(2015年9期)2015-03-01
