
人α-突触核蛋白A30P突变对小鼠成体大脑吻端迁移流中神经发生的抑制作用及其机制
2013-09-19 05:47李玲玲孙艺学王爱兵李子义张学明
吉林大学学报(医学版) 2013年6期
李玲玲,孙艺学,王爱兵,李子义,张学明
(1.吉林省前卫医院内二科,吉林 长春 130012;2.吉林大学动物医学学院 动物胚胎工程吉林省重点实验室,吉林 长春 130062)
成体大脑中的神经发生主要存在于脑室下区(subventricular zone,SVZ)和海马齿状回颗粒下区(subgranular zone,SGZ)。SVZ中的神经干细胞增殖后沿吻端迁移流(rostral migratory stream,RMS)分化并迁移至溴球(olfactory bulb,OB),然后整合入OB突触球层中的神经网络而发挥功能[1]。人退行性神经性疾病如帕金森病和阿尔茨海默病等均与OB功能严重障碍有关联[2]。α-突触核蛋白(α-synuclein,SNCA)表达于OB中的受体神经元,提示这些细胞及来自SVZ的神经细胞有可能受到SNCA突变的影响[3]。SNCA是一个由140个残基组成的神经元蛋白。在生理条件下,大部分存在于神经突触前末梢,靠近突触小泡。研究[4]证实:SNCA聚集物存在于许多病理进程中,如帕金森病、阿尔茨海默病和路易体痴呆等。一些家族性帕金森病患者的SNCA基因中第209位核苷酸发生G→A突变,使氨基酸序列中第53位丙氨酸(Ala)变成苏氨酸(Thr),记为 A53T[5];第88位碱基发生G→C突变,导致氨基酸序列第30位丙氨酸变成脯氨酸(Pro),记为A30P[6];或第188位碱基发生G→A突变,使氨基酸序列中第46位谷氨酸(Glu,E)变成赖氨酸(Lys,K),记为E46K[7]。有关SNCA这3种点突变对成体大脑神经发生的影响尚不清楚。本研究探讨了SNCA A30P突变对小鼠大脑RMS中神经细胞增殖的影响。
1 材料与方法
1.1 实验动物、主要试剂和仪器 采用英国牛津大学Francis G.Szele博士实验室提供12只1~2月龄C57BL/6正常小鼠,雌雄各半。将4只C57BL/6正常小鼠、4只内源性SNCA敲除小鼠(SNCA-/-)、4只内源性SNCA 敲除+ 人SNCA A30P敲入小鼠(A30PSNCA-/-)分别作为正常对照组、SNCA-/-组和 A30PSNCA-/-组。一抗磷酸化组蛋白 3(phosphohistone 3,PH3,
Millipore公司)和双皮质素(doublecortin,Dcx,Santa Cruz公司),二抗Cy3和Alexa Fluro 488(Jackson Immunoresearch公司),4′,6-二脒基-2-苯 基 吲 哚(4′, 6-diamidino-2-phenylindole dihydrochloride,DAPI,Sigma-Aldrich公司),荧光淬灭剂(武汉博士德公司);荧光显微镜(日本尼康公司),冰冻切片机(CM1900,德国莱卡公司)。
1.2 脑组织冠状冰冻切片的制备 深度麻醉各组小鼠,剪开胸廓,暴露搏动的心脏,心脏左心室穿针,剪开右心耳,缓慢注入4℃生理盐水,再注入4℃、4%多聚甲醛固定,枕骨大孔处用剪刀横断,剪开顶骨,用止血钳捌断顶骨,取出完整大脑。置于4%多聚甲醛中4℃下固定过夜,再用30%蔗糖平衡。-80℃保存,冰冻切片厚30μm,置于防冻液中于-20℃保存备用。
1.3 荧光免疫组织化学法染色 每只小鼠选取RMS切片的前、中和后部位各3张,用PBS洗涤3×10min,50mmol·L-1甘氨酸处理15min后PBS洗涤3×10min;换PBS(含有10%驴血清和0.1%Triton X-100的PBS溶液),室温孵育1h;用PBS+稀释一抗PH3(兔抗,1∶500)和Dcx(山羊抗,1∶100)4℃孵育过夜;PBS洗涤3×10min,PBS+稀释二抗Cy3(驴抗兔,1∶500)和Alexa Fluro 488(驴抗山羊,1∶500)孵育1h;PBS洗涤3×10min,用40g·L-1DAPI原液1∶1 000溶于PBS进行核染色10min;PBS洗涤3×10min,自然风干后用荧光淬灭剂封片,自然避光凉干后荧光显微镜或激光扫描共聚焦显微镜下观察、拍照,统计PH3阳性(PH3+)细胞数,比较各组中DAPI染色细胞的密度和Dcx荧光强度。此外,在伯特兰透镜(bertrand lens,×1)下拍照,观察各组小鼠脑组织RMS的形态学。
1.4 统计学分析 采用SPSS 13.0统计软件进行数据分析。各组PH3+细胞数以±s表示,组间比较采用t检验。
2 结 果
2.1 各组小鼠脑组织RMS形态学 3组小鼠切片标本中RMS部位切片的形状、大小和细胞密度等未见明显变化,箭头所示处为RMS,正常对照组、SNCA-/-和 A30PSNCA-/-组小鼠脑组织 RMS的形状、大小及其细胞密度眼观均无明显变化。见图1。
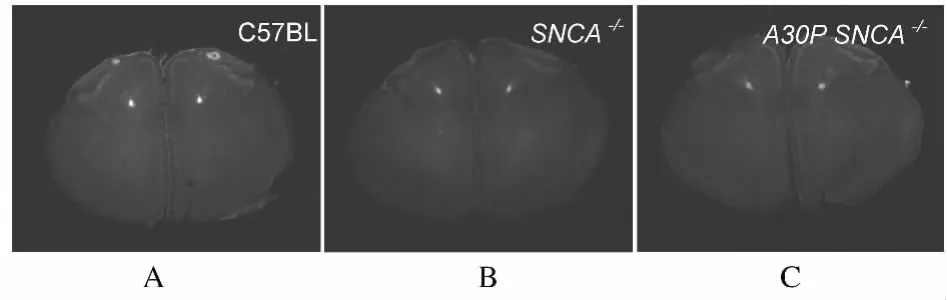
图1 各组小鼠脑组织切片RMS的形态学Fig.1 Morphology of RMS of mouse brain section in various groups
2.2 各组小鼠脑组织RMS中的细胞增殖 荧光免疫组织化学分析,RMS中增殖性神经细胞呈亮黄色。正常对照组、SNCA-/-和 A30PSNCA-/-组小鼠RMS组织中PH3+细胞数分别为(7.4±1.9)、(5.0±1.6)和(4.8±1.7)个;与正常对照组比较,SNCA-/-组和 A30PSNCA-/-组小鼠RMS中PH3+细胞数量明显减少(P<0.05)。DAPI染色,RMS的细胞密度和荧光强度未见明显变化。见图2(插页四)。
2.3 各组小鼠脑组织RMS中神经母细胞的迁移
Dcx荧光免疫染色,与正常对照组比较,SNCA-/-组小鼠神经母细胞的荧光强度增强,A30PSNCA-/-组小鼠的Dcx阳性(DCX+)神经母细胞荧光强度则轻微增强。见图2(插页四)。
3 讨 论
Polymeropoulos等[5]于1997年首次描述了家族性帕金森患者SNCA A53T突变;随后Kruger等[6]于1998年发现了人SNCA A30P突变;2004年,Zarranz等[7]报道数例家族性帕金森患者中存在SNCA E46K突变。但目前有关3种点突变可能引发人退行性神经疾病的机制尚不清楚。Winner等[8]用转基因小鼠研究了SNCA A53T突变对大脑神经发生的影响,发现由血小板衍生生长因子(PDGF)启动子驱动的SNCA A53T突变能减弱SVZ中的神经发生且上调凋亡。但第53位的苏氨酸(T)是小鼠体内一种正常氨基酸,因此本文作者认为:利用SNCA A53T转基因小鼠研究人SNCA A53T突变对小鼠大脑神经发生的影响是不可取的。研究者[9-10]采用四环素条件性转基因小鼠系统研究人SNCA A30P突变对小鼠大脑神经性发生的影响发现:人A30P突变能诱发小鼠OB中新生神经元的丢失。但这种体系未敲除小鼠内源性SNCA,因此转基因的表达有可能被内源性SNCA屏蔽。本研究采用先敲除C57BL小鼠内源性SNCA,再导入人SNCA A30P的策略,结果更为可靠。至于SNCA E46K突变,目前尚未有转基因鼠可用。
从形态学观察来看,敲除内源性SNCA或敲除后再导入人SNCA A30P突变对小鼠大脑RMS部位切片的形态、大小和结构影响甚微。组蛋白3(histone,H3)的Ser10、Ser28和Thr11均会发生磷酸化修饰,这些磷酸化和有丝分裂或减数分裂过程中的染色质浓缩有密切关联,因此PH3常被用以检测细胞增殖情况。本实验中PH3漂浮切片免疫组织化学结果显示:内源性SNCA敲除或SNCA敲除+人SNCA A30P敲入对RMS中的神经细胞增殖情况有较为明显的抑制作用,提示正常SNCA的表达对成体大脑神经发生可能是必要的,同时也说明人SNCA A30P突变可能减弱成体大脑神经发生。Dcx的染色结果提示:敲除内源性SNCA或再导入人SNCA A30P,虽然减弱了小鼠大脑神经增殖活动,但对该处已增殖神经元向神经母细胞的分化以及分化后神经母细胞向OB的迁移可能有一定促进作用。这一点看似矛盾,但也可从帕金森病遗传学调查中得到一定解释。研究[11-12]发现:多数帕金森病患者均为不明原因的散发性病例,家族性病例则一般由单基因突变,特别是SNCA的3个误义突变(A53T、A30P和E46K)及其拷贝数突变所引起。换言之,SNCA及其突变只是一部分家族性帕金森病发生的原因,而另一部分帕金森病的发生可能与SNCA突变关系不大。因此,在临床上仍需进一步筛查帕金森病(包括散发性和家族性的)发生的关键基因。
[致谢:感谢国家留学基金委对本项目的部分资助,特别感谢英国牛津大学Francis G.Szele博士及其实验室内原成员Yongsoo Kim博士、Isabelle Comte博士和Christopher Young博士等对本研究的支持。]
[1]孙艺学,王爱兵,李子义,等 .成体大脑神经发生研究进展 [J].吉林大学学报:医学版,2013,39(5):1076-1080.
[2]Kovacs T.Mechanisms of olfactory dysfunction in aging and neurodegenerative disorders [J].Ageing Res Rev,2004,3(2):215-232.
[3]Duda JE,Shah U,Arnold SE,et al.The expression of alpha-,beta-,and gamma-synucleins in olfactory mucosa from patients with and without neurodegenerative diseases[J].Exp Neurol,1999,160(2):515-522.
[4]Kohl Z,Winner B,Ubhi K,et al.Fluoxetine rescues impaired hippocampal neurogenesis in a transgenic A53T synuclein mouse model[J].Eur J Neurosci,2012,35(1):10-19.
[5]Polymeropoulos MH,Lavedan C,Leroy E,et al.Mutation in the alpha-synuclein gene identified in families with Parkinson's disease[J].Science,1997,276(5321):2045-2047.
[6]Kruger R,Kuhn W,Muller T,et al.Ala30Pro mutation in the gene encoding alpha-synuclein in Parkinson's disease[J].Nat Genet,1998,18(2):106-108.
[7]Zarranz JJ,Alegre J,Gomez-Esteban JC,et al.The new mutation,E46K,of alpha-synuclein causes Parkinson and Lewy body dementia[J].Ann Neurol,2004,55(2):164-173.
[8]Winner B,Rockenstein E,Lie DC,et al.Mutant alphasynuclein exacerbates age-related decrease of neurogenesis[J].Neurobiol Aging,2008,29(6):913-925.
[9]Marxreiter F,Nuber S,Kandasamy M,et al.Changes in adult olfactory bulb neurogenesis in mice expressing the A30P mutant form of alpha-synuclein[J].Eur J Neurosci,2009,29(5):879-890.
[10]Nuber S, Petrasch-Parwez E, Winner B, et al.Neurodegeneration and motor dysfunction in a conditional model of Parkinson’s disease [J].J Neurosci,2008,28(10):2471-2484.
[11]Chartier-Harlin MC, Kachergus J, Roumier C,et al.Alpha-synuclein locus duplication as a cause of familial Parkinson’s disease[J].Lancet,2004,364(9440):1167-1169.
[12]Singleton AB,Farrer M,Johnson J,et al.Alpha-synuclein locus triplication causes Parkinson’s disease [J].Science,2003,302(5646):841.
猜你喜欢
中国临床医学影像杂志(2022年6期)2022-07-26
保健医苑(2022年6期)2022-07-08
昆明医科大学学报(2021年8期)2021-08-13
国际放射医学核医学杂志(2021年10期)2021-02-28
科学(2020年3期)2020-11-26
心理学报(2020年7期)2020-07-13
中国临床医学影像杂志(2019年1期)2019-04-25
活力(2019年22期)2019-03-16
中国生殖健康(2019年9期)2019-01-07
现代养生·下半月(2015年6期)2015-09-07
