
人血管内皮生长因子的生物信息学分析
2013-09-10 05:36广东省信宜人民医院心内科广东信宜525300
吉林医学 2013年36期
梁 丕(广东省信宜人民医院心内科,广东 信宜 525300)
血管内皮生长因子(Vascular endothelial growth factor,VEGF),又称血管通透因子(Vascular permeability factor,VPF)或血管调理素(Vasculotropin),是1989年Ferrara等从牛垂体滤泡星状细胞培养液中分离纯化的糖类蛋白质,后来在鼠垂体前叶肿瘤细胞系、鼠神经胶质瘤细胞、豚鼠瘤、人单核细胞等细胞培养液中也纯化出了VEGF蛋白[1]。VEGF是一个具有高度保守性,由两个相同亚基(Mr 23 000)以二硫键交联结合成的二聚体糖蛋白,主要作用于血管内皮细胞[2-3]。VEGF由某些肿瘤细胞分泌,通过与血管内皮上的相应受体结合促进内皮细胞增殖,同时可增加血管通透性使内皮细胞迁移,诱导肿瘤血管生成,维持肿瘤的继续生长,是目前发现的最强烈的血管生成因子,与肿瘤及非肿瘤的血管新生病理过程中密切相关,其与转化生长因子-β(TGF-β)、血小板源性生长因子(PDGF)、NO及一些重金属离子有关,还受缺血和缺氧环境的调节[4]。目前VEGF基因已先后从人、野猪、绵羊等多种动物中克隆出来,并通过分子遗传学方法证明了该基因在血管生成、重构、病变等过程中的重要作用,因此对VEGF的研究成为当前的热点课题。本文利用生物信息学工具和方法,以人VEGF分析为重点,对动物VEGF的核苷酸和氨基酸序列的特征,及其编码蛋白的理化性质、分子结构、生化功能和系统演化等进行预测分析,以期为深入探讨VEGF蛋白的酶学特性和血管生成的分子机制提供理论依据。
1 临床资料
VEGF的核苷酸和氨基酸序列来源于NCBI数据库,其中人VEGF的登录号依次为:GenBank Accession No.AY047581,GenPept accession no.AAK95847。
利用Vector NTI Suit 8、Clustal X、MEGA4.1β、Accelrys ViewerLite 5.0软件和www.ncbi.nlm.nih.gov、www.expasy.org等网站提供的各种在线生物信息学工具进行预测和分析。核酸及氨基酸序列的分子结构和理化性质分析、开放阅读框的查找和翻译使用Vector NTI Suit 8和Protparam软件完成;核酸及氨基酸序列的同源性比对使用Clustal X软件和NCBI上的Blast在线工具进行;分子系统发生树使用MEGA3的Phylogeny中的Neighbor-Joining方法构建;蛋白质的功能结构域和三级建模分别用CDD和SWISSMODEL分析。
2 结果
2.1 人VEGF基因的核酸及其编码蛋白的序列特征和理化性质:NCBI上登录的人VEGF核苷酸序列全长640 bp,包括3'-UTR(Untranslated Regions,非翻译区)、ORF(Open Reading Frame,开放阅读框)和5'-UTR序列,且编码区的起始密码子为ATG,终止密码子为TGA。用Protparam预测VEGF编码蛋白的理化性质,推测该蛋白的分子式为C960H1504N288O281S23,分子量为22.3136 kDa,等电点pI=7.88,摩尔消光系数为24 950,半衰期为30 h,脂肪酸系数60.73,总的亲水性平均系数(Grand average of hydropathicity,GRAVY)为-0.704,推论该蛋白属于疏水性蛋白,而不稳定参数为56.69,故属不稳定蛋白。该基因相应的氨基酸残基数为191,所含氨基酸组成见表1,含Cys和Glu最多,不含Pyl和Sec。
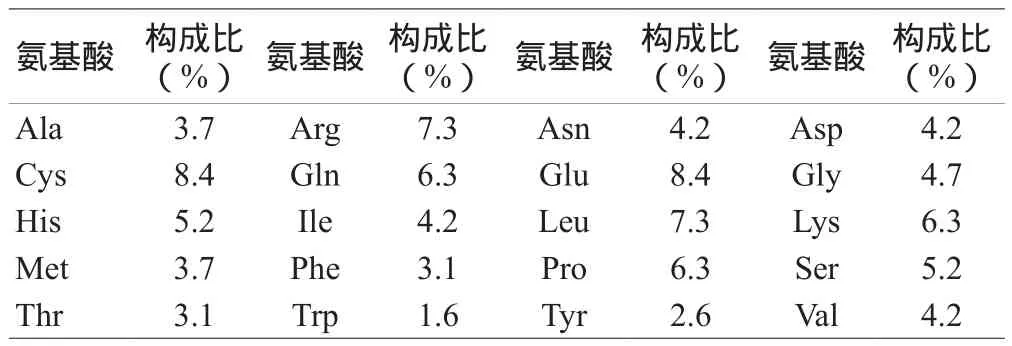
表1 人VEGF蛋白的氨基酸组成
2.2 人VEGF蛋白的氨基酸序列的比对分析:选取人等7条动物VEGF氨基酸序列,在Vector NTI 8软件中进行多重比对分析。分析结果用不同颜色显示,见图1,颜色越深的表示同源性越高,最深的区域则表示可能存在的重要功能域。分析结果显示动物VEGF具有很高的同源性,一致性达到43.5%,可见VEGF编码区高度保守。而其中仅原鸡的变异性较大,则提示禽类与哺乳动物的VEGF之间存在差异,这一方面说明VEGF在物种进化过程具有重要意义,另一方面也推测这可能是由于动物血管生成的机制存在很强的相似性,而作为与血管发生密切相关的VEGF蛋白必须保持足够的遗传稳定性和演化趋同性。

图1 来自不同动物VEGF氨基酸序列多重比对
2.3 人VEGF蛋白的氨基酸序列的分子系统进化树分析:将7条动物VEGF蛋白氨基酸序列在Clustal X软件中进行完全比对后,在MEGA4.1软件中依据NJ(Neighbor-Joining)方案进行构建分子进化树[5-6]。获得VEGF氨基酸序列的分子进化树分析结果见图2。7种动物的VEGF氨基酸序列聚成4支:人与野猪聚成第一支;绵羊和藏羚羊聚成第二支;鼹鼠和小鼠聚成第三支,而原鸡则独立一支。该进化树很好地反映出了这7种不同动物之间的亲缘关系,如人与猪同属较高等的哺乳动物,聚为一支,绵羊与藏羚羊同属羊亚科也聚为一支,鼹鼠和小鼠为鼠总科的啮齿类动物,原鸡则因其属非哺乳的禽类而单独成支。由此可见,依据氨基酸序列构建的分子进化树能在一定程度上反映不同动物之间的系统分类学关系和自然演化进程,其结果对于明确判断物种间的亲缘关系能提供一定的参考。
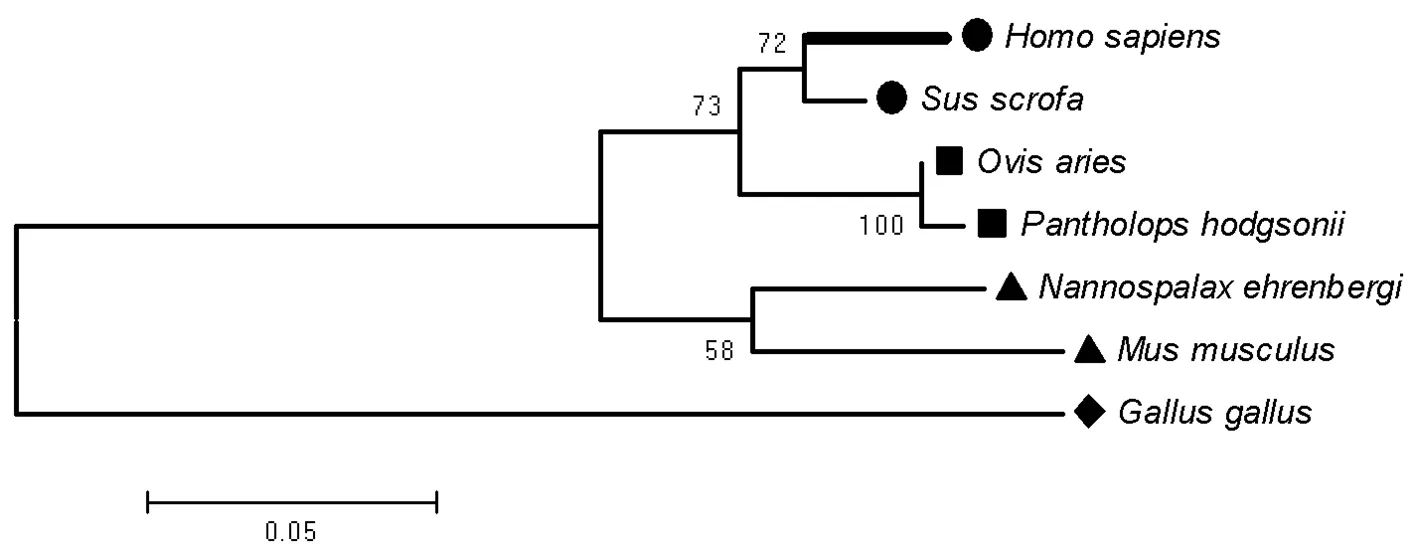
图2 基于MEGA4.1中NJ方法的人和其他物种的VEGF分子进化树聚类分析
2.4 人VEGF编码蛋白的功能结构域和三维建模分析:蛋白质由多个基序组成,且基序相应的氨基酸区段行使特异的生化功能,同时也蕴含着各自的遗传进化信息。利用CDD在线工具分析人VEGF氨基酸序列的功能结构域,即去掉信号肽,剩下的成熟肽氨基酸序列包含一个PDGF超家族功能域,见图3[7]。表明人VEGF编码蛋白属于血小板衍生因子(Platelet-derived growth factor,PDGF)超家族,同样属于一种重要的促有丝分裂因子,具有刺激特定细胞群分裂增殖的能力,在血管生成和重构机制中具有关键作用。将这条人VEGF氨基酸序列上传至SWISS-MODEL服务器中进行基于同源性的三维建模,然后在ViewerLite 4.2软件中进行序列编辑,获得人VEGF的三级结构模型,见图4[8-9]。
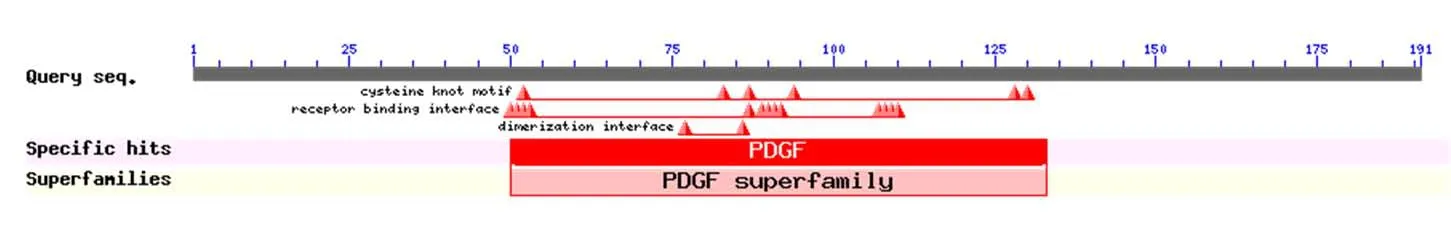
图3 人VEGF的保守结构域分析

图4 人VEGF基因编码蛋白的三维结构预测
3 讨论
VEGF作为血管生成和重构的关键因子而倍受关注,依托于生物学实验数据与现代计算技术相结合的生物信息学已经成为后基因组时代的重要方法和前沿领域[10]。本文应用生物信息学方法对人VEGF基因的核苷酸及其编码蛋白质序列进行了比对、分析和建模,进而对其分子结构和生化功能进行了预测分析,结果表明该基因的核苷酸序列包括3'-UTR、ORF和5'-UTR,推测其编码蛋白分子式为C960H1504N288O281S23,是含有PDGF重要功能域亲水性不稳定蛋白质,其在动物系统进化过程中可能具有重要作用,且三维结构建模成功。本研究结果可为深入开展人VEGF蛋白的生化特性和血管生成的分子机制研究提供重要理论参考,有利于推动桑等肿瘤学和心血管病学的发展。
[1] Senger DR,Water L,Brown LF,et al.Vasular permeability factor(VPF,VEGF) in tumor biology[J].Cancer Metastasis Rev,1993,12(3):303.
[2] Ferrara N,Henzel WJ.Pituitary follicular cells secrete a novel heparin binding growth factor specific for vascular endothelial cells[J].Biochem Biophys Res,1989,161(6):851.
[3] Myoken Y,Kayada Y,Okamoto T,et al.Vascular endothelial cell growth factor(VEGF) produced by A-431 human epidermoid carcinoma cells and identification of VEGF membrane binding sites[J].Proc Natl Acad Sci USA,1991,88(30):5819.
[4] 王彦敏.血管生成因子VEGF研究进展[J].河北医药,2010,32(11):1456.
[5] Thompson JD,Gibson TJ,Plewniaki F,et al.The CLUSTALX windows interface:flexible strategies for multiple sequence alignment aided by quality analysis tools[J].Nucleic Acids Res,1997,18(25):4876.
[6] Saito N,Nei M.The neighbor joining method:a new method for reconstructing phylogenetic trees[J].Mol Biol Evol,1987,18(4):406.
[7] Marchler A,Bryant SH.CD-Search:protein domain annotations on the fly[J].Nucleic Acids Res,2004,32(3):327.
[8] Schwede T,Kopp J,Guex N,et al.SWISS-MODEL:an automated protein homology-modeling server[J].Nucleic Acids Research,2003,31(13):3381.
[9] Guex N,Peitsch MC.SWISS-MODEL and the Swiss-PdbViewer:An environment for comparative protein modeling[J].Elec trophoresis,1997,18(15):2714.
[10] 蔡源源,李光早.血管内皮生长因子的调控及其作用研究进展[J].组织工程与重建外科杂志,2011,7(1):51.
猜你喜欢
天津市教科院学报(2021年5期)2021-11-10
生物学通报(2021年9期)2021-07-01
中国洗涤用品工业(2019年4期)2019-05-11
中成药(2018年1期)2018-02-02
中成药(2017年3期)2017-05-17
中国医学创新(2017年7期)2017-03-31
江苏农业科学(2016年8期)2017-02-15
中国中西医结合皮肤性病学杂志(2016年4期)2016-07-18
中国医药生物技术(2015年4期)2015-12-26
中国现代医学杂志(2015年26期)2015-12-23
