
玉米花生间作对玉米光合特性及产量形成的影响
2013-09-07 02:53焦念元宁堂原杨萌珂付国占徐国伟李增嘉
生态学报 2013年14期
焦念元,宁堂原,杨萌珂,付国占,尹 飞,徐国伟,李增嘉
(1.河南科技大学农学院,洛阳 471003;2.山东农业大学农学院/作物生物学国家重点实验室,泰安 271018)
合理的间套作能高效利用光、热、肥、水等自然资源,降低风险,减少杂草竞争,提高单位耕地面积的产量[1-2],在我国以及世界农业中发挥着重要作用。随着世界人口急剧增长和耕地面积的减少,不断满足世界粮食增长的需求已刻不容缓,间套作多熟种植越来越受到世界各国农业生产的重视。在纵多间套作种植体系中,玉米处于非常重要的地位[2]。已有研究表明,在玉米菜豆[3]、玉米蚕豆[4]、玉米大豆[5]、玉米花生[6-8]等间作体系中,间作玉米产量均明显高于单作,表现出明显的间作产量优势。
在玉米与其他作物间套作体系中,全田群体高矮相错,形成伞状群体结构,改变了玉米生育后期单一群体的平面受光状态,受光条件得到改善,提高了功能叶片的叶绿素含量、蒸腾速率、气孔导度和光合速率[8-9]。玉米花生间作是东亚和非洲常见的种植体系,近年来在我国四川、广东以及黄淮海等地区发展较快,种植面积不断扩大,间作提高玉米对强光利用[6-7],表现出显著的间作产量优势[8]。间作玉米提高对强光的利用,是光能吸收传递效率的提高,还是光合作用CO2的同化过程增强引起?还需进一步研究,同时间作玉米产量形成特点发生哪些适应性变化?对此,本试验以玉米花生2∶4间作模式为研究对象,主要研究了间作玉米光-光响应曲线和光-CO2响应曲线特点、叶绿素含量与构成、干物质积累、灌浆速率及间作玉米偏土地当量比(PLERM),探讨间作玉米光合特性,进一步揭示玉米间作产量优势的光合机理,为协调玉米花生间作后期种间光竞争,实现其高产高效提供理论依据。
1 材料与方法
1.1 供试材料与试验设计
试验于2010—2011年在河南科技大学试验农场(33°35'—35°05'N,111°8'—112°59'E)进行。试验地属温带半湿润半干旱大陆性季风气候,年均气温12.1—14.6℃,年均降水量600 mm,年均辐射量491.5 kJ/cm2,年日照2300—2600 h,无霜期215—219 d。试验地土壤为黄潮土,质地中壤,0—20 cm耕层含速效氮80.09 mg/kg,速效磷 3.31 mg/kg,有机质 14.5g/kg,pH 值 7.08。
2010年选用玉米(Zea mays L.)郑单958、登海661,花生(Arachis hypogaea L.)花育16为试验材料。设郑单958单作、登海661单作、郑单958花生间作和登海661花生间作,共4个处理,各处理重复3次,共12个小区,每个小区宽6 m,包含3个带宽,长10 m,面积60 m2,随机排列,南北行向种植。单作玉米行距60 cm,株距25 cm,密度67500株/hm2;间作体系2∶4(2行玉米4行花生)间作模式,玉米宽窄行种植,宽行行距160 cm,窄行行距40 cm,株距20 cm,花生播种于宽行中,行距30 cm,株距20 cm,每穴2粒;玉米花生间距35 cm。基施氮肥75 kg N/hm2,磷肥180 kg P2O5/hm2,在玉米大口期追施150 kg N/hm2。6月13日播种,9月30日收获。
2011年玉米只选郑单958,6月15日播种,10月7日收获,其他同2010年。
1.2 测定项目与方法
1.2.1 干物质
分别于玉米苗期、拔节期、大喇叭口期、吐丝期、授粉后10 d、授粉后20 d、授粉后30 d和收获期取样,分茎、叶、穗轴、籽粒四部分,105℃下杀青30 min,75℃烘干至恒重称重。
1.2.2 灌浆速率
在吐丝期,选择吐丝日期相同的植株进行标记,分别于授粉后10 d、20 d、30 d、40 d和收获期取3个果穗,105℃下杀青30 min,75℃烘干至恒重,测定穗干重。
1.2.3 收获测产
在玉米成熟期,各处理均取具有代表性的4m双行果穗和植株,测定其风干质量,计算产量。
1.2.4 光合参数测定
用英国PP-systems公司生产的CIRAS-2型便携式光合系统在玉米灌浆期和乳熟期测定穗位叶9:00—11:00时净光合速率;在乳熟期9:00—11:00时,采用自动可调光源,分别测定自然CO2浓度下光强为1800、1500、1200、1000、800、600、500、400、300、200、150、100、50μmol·m-2·s-1的光 合 速 率 (Pn);在光 强1200μmol·m-2·s-1下,分别测定 CO2浓度为 400、350、300、250、200、150、100、50、400、400、600、800、1000、1200 μmol/mol时的光合速率(Pn),应用photosyn assistant软件分析计算光补偿点(LCP)、光饱和点(LSP)、光饱和时净光合速率(LSPn)、表观量子效率(AQY)、CO2饱和点(CSP)、羧化效率(CE)。
1.2.5 叶绿素荧光参数测定
在玉米乳熟期用英国Hansatech公司生产的FMS2脉冲调制式荧光仪测定光适应下最大荧光(Fm`)、光适应下的稳态荧光(Fs)和暗适应30min后的最大荧光(Fm)、初始荧光(Fo)。计算PSⅡ的最大光化学效率(Fv/Fm)、实际光化学效率(ΦPSⅡ)和光化学猝灭系数qP公式分别为:
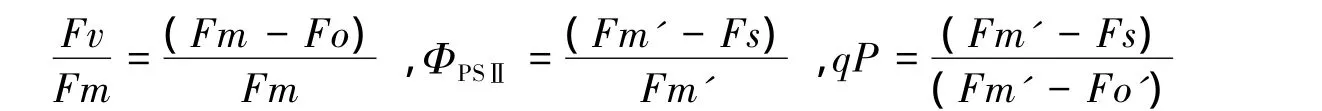
1.2.6 叶绿素含量
采用Arnon法[9],在乳熟期取玉米的穗位叶测定。
1.2.7 偏土地当量比 PLER-M=Yim/Ymm
式中,Yim和Ymm分别表示间作玉米和单作玉米产量。PLER-M>F(间作体系中玉米所占面积比例)为间作产量优势,PLER-M<F为间作产量劣势。本试验中F为0.4。

表1 间作对玉米穗位叶叶绿素含量的影响Table1 Effect of intercropping on chlorophyll content of maize ear leaves
1.3 数据统计分析 采用DPS7.5软件对数据进行差异显著性检验。
2 结果与分析
2.1 玉米花生间作对玉米光合特性影响
2.1.1 间作玉米功能叶片的叶绿素含量及构成
叶绿素在植株体内负责光能的吸收、传递和转化,类胡萝卜素具有光能捕获和光破坏防御两大功能,在光合作用中起着非常重要的作用[10]。由表1可以看出,与单作玉米相比,间作玉米穗位叶的叶绿素a含量、类胡萝卜素含量、叶绿素a/b和类胡萝卜素/叶绿素分别增加了3.69%、6.52%、2.67%和4.17%,差异均达到极显著水平。这说明间作玉米通过提高叶绿素a含量和叶绿素a/b促进光能的吸收,同时增加类胡萝卜素含量和类胡萝卜素/叶绿素增强光破坏防御能力,提高光合速率。
2.1.2 间作玉米功能叶片的荧光特性
Fo是植物叶片暗适应后光合系统PSⅡ中心完全开放时的荧光强度,反映了PSⅡ天线色素受激发后的电子密度;Fv是植物在暗适应过程中的最大可变荧光强度,反映了QA的还原情况[11-12];Fv/Fm是PSⅡ最大的(潜在)光化学量子效率,反映开放的PSⅡ反应中心的能量捕获效率;ΦPSⅡ是作用光存在时PSⅡ实际的光化学量子效率,反映了被用于光化学途径激发能占进入PSⅡ总激发能的比例,植物光合能力的一个重要指标;qP为光化学淬灭系数,反映PSⅡ天线色素吸收的光能用于光化学电子传递的份额。由表2可得,与单作玉米相比,间作降低了玉米功能叶片的初始荧光、可变荧光和实际光化学效率,但差异均不显著;间作对玉米功能叶片的最大光化学效率和光化学猝灭系数影响不明显。这表明,间作玉米光合速率的提高并不是由于其功能叶片对光能传递、转化效率提高而引起的。
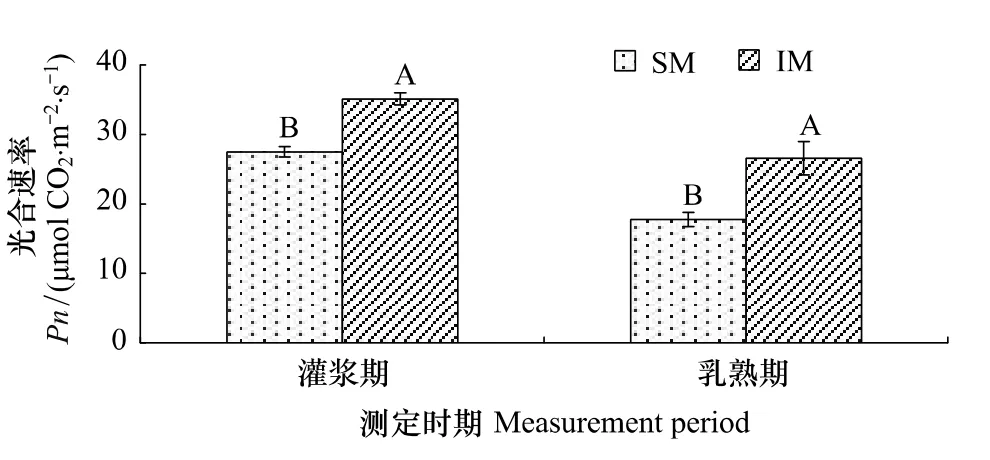
图1 间作对玉米功能叶片光合速率的影响Fig.1 Effect of intercropping on photosynthetic rate of maize leaves
2.1.3 间作玉米功能叶的光合特性
图1表明,间作明显提高了玉米功能叶片的净光合速率,在灌浆期和乳熟期分别提高了27.7%和49.8%,差异均达到显著水平,由灌浆期到乳熟期,单作玉米和间作玉米光合速率均降低,其降低幅度分别为35.5%和24.3%。这表明玉米花生间作显著提高了玉米净光合速率,减缓玉米叶片衰老,延长其高光合功能期。
由表3可以看出,与单作玉米相比,间作提高了玉米功能叶片的光补偿点、光饱和点、光饱和时的最大净光合速率、表观量子效率和羧化效率,分别提高了26.7%、53.1%、33.1%、61.3%、8.5%和22.5%,差异达到显著或极显著水平,极显著降低了 CO2补偿点。这表明间作玉米光合速率的提高是由于提高了其对强光的利用能力和CO2的羧化能力。

表3 间作对玉米光合参数的影响Table3 Effect of intercropping on photosynthetic parameters of maize leaves
2.2 玉米花生间作对玉米产量形成的影响
2.2.1 间作玉米单株干物质
单株干物质是衡量玉米生长状况的重要指标之一。从图2可得,在玉米大喇叭口期之前,间作玉米的单株干物质质量与单作相差不明显,之后间作玉米单株干物质质量逐渐高于单作玉米,平均高出7.64%—30.9%。尤其2011年,生育后期的间作玉米比单作玉米高出20.9%—30.9%,差异达到显著水平。从单株茎叶质量来看,间作玉米与单作玉米差异不明显。
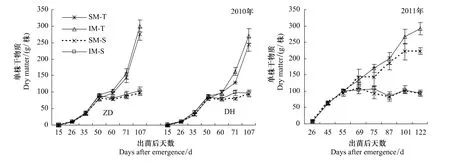
图2 间作玉米单株干物质积累Fig.2 Dry matter accumulation of intercropped maize
2.2.2 间作玉米灌浆速率
玉米穗重变化是衡量玉米灌浆速率指标之一。从图3可以看出,自玉米授粉之后,间作玉米单穗质量逐渐高于单作玉米,随灌浆进程,其差异越来越明显,授粉20 d后,间作玉米的单穗质量比单作玉米高出8.8%—32.8%,在收获期差异达到显著水平。这表明,间作玉米与单作玉米单株质量差异主要来源生殖器官即单穗籽粒质量的提高,其营养器官即茎叶质量差异不明显。
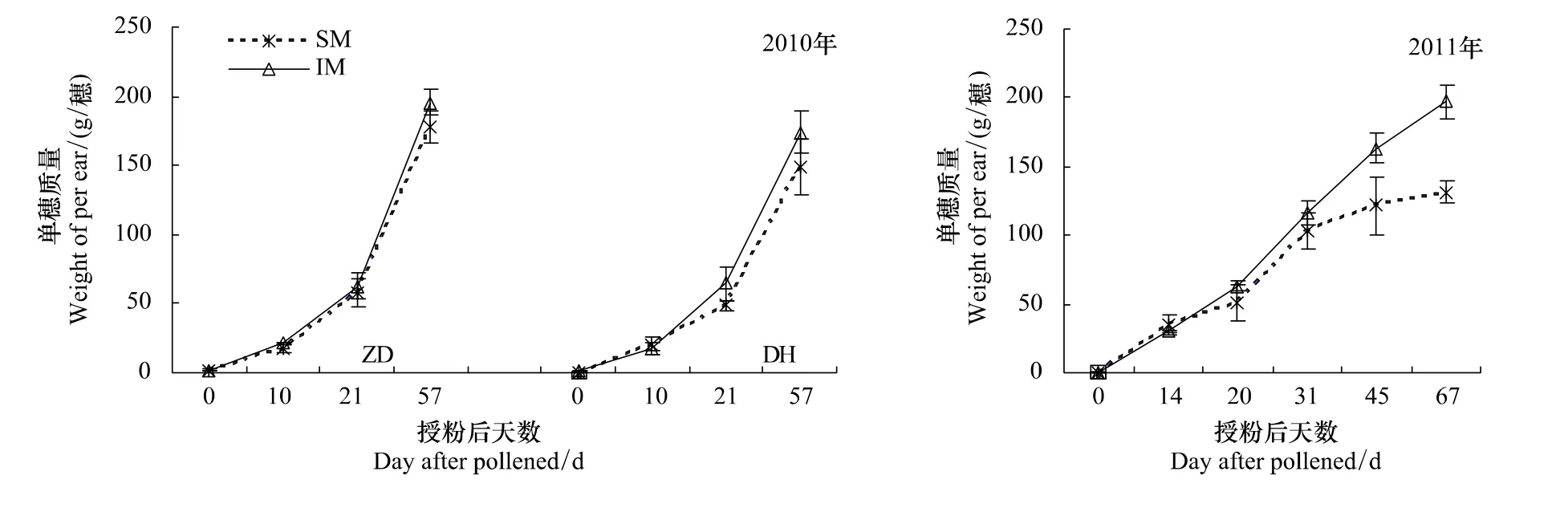
图3 间作玉米灌浆速率Fig.3 Filling rate of intercropped maize
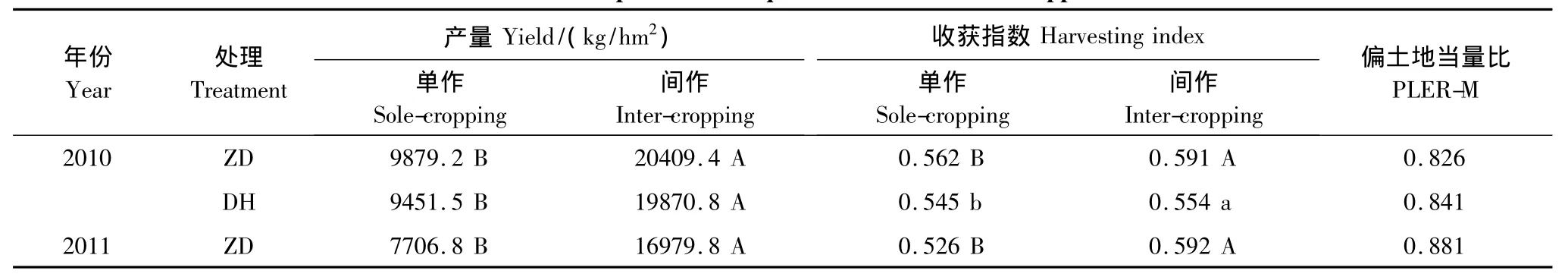
表4 间作玉米产量与偏土地当量比Table4 Yield and partial land equivalent ratios of intercropped maize
2.2.3 间作玉米产量与偏土地当量比
从表4可以看出,与单作相比,间作极显著提高了玉米产量,间作玉米的收获指数显著或极显著高于单作玉米,其偏土地当量比(PLER-M)比其所占土地面积比(F=0.4)高出106.6%—120.3%。这表明在玉米花生间作体系中,促进了光合物质向籽粒的运输、分配,表现出明显的间作产量优势。郑单958的PLER-M低于登海661,但是,郑单958收获指数和产量均高于登海661。
3 讨论
干物质是作物产量形成的基础。本研究发现,间作玉米单株干物质仅在生育后期明显高于单作玉米,并且其茎叶等营养器官干物质在各个生育时期均差异不大,但其生殖器官——单穗质量差异明显。这主要因为间套作有利于改善高位作物生育后期冠层光照条件[6,13-14],提高了功能叶片净光合速率,促进籽粒灌浆和光合物质向籽粒运输与分配,因此,间作玉米具有高的收获指数,PLER-M比间作玉米所占间作体系中面积比例(F=0.4)高出106.6%—120.3%,表现出明显的间作产量优势。这与多数研究具有一致的结论,与其他作物间作时,玉米具有明显间作产量优势[3-7]。
叶绿素在植株体内负责光能的吸收、传递和转化,类胡萝卜素则行使光能捕获和光破坏防御两大功能,它们在光合作用中起着非常重要的作用,其含量及组成受光照条件的影响[15]。本研究表明,在玉米花生间作体系中,间作改善玉米生育后期光照条件,不仅提高叶绿素含量,还改变了叶绿素构成,主要表现在叶绿素a和类胡萝卜素含量均显著增加,叶绿素a/b和类胡萝卜素/叶绿素的比值均显著增加。因此,在强光下,间作玉米通过提高叶绿素a、类胡萝卜素含量来增加光反应中心,促进将吸收较多光能进行光反应,提高净光合速率。类胡萝卜素具有光能捕获和光破坏防御两大功能,其含量增加有助于提高光破坏防御能力,延缓衰老。
光环境影响着植物的光合特性[16],在玉米花生间作体系中,间作玉米功能叶片接受的光照强度高于单作玉米,其净光合速率提高了27.7%—49.8%,光饱和点、光饱和时最大光合速率、羧化效率显著提高,但是间作玉米功能叶片PSⅡ的最大光化学量子效率和实际光化学量子效率没有提高,也就说是PSⅡ反应中心的被用于光化学途径激发能占进入PSⅡ总激发能的比例并没有提高。间作玉米光合效率的提高是由于呼吸速率提高为固定CO2提供能量,降低CO2补偿点,极显著提高了羧化效率、表观量子效率,提高了CO2的固定能力,这与朱文旭[17]的研究具有相同结论,间作提高高位作物的羧化能力。这可能与光照条件影响叶绿体超微结构[18]、光合酶活性和光合基因表达[19]有关,但对间作条件下作物功能叶的叶绿体超微结构、光合酶活性和光合基因表达发生发些适应性变化还需进一步研究。
4 结论
在玉米花生间作体系中,间作玉米具有明显间作产量优势,是由于间作增强了玉米对强光利用能力,提高了玉米生育后期功能叶片的净光合速率,促进了光合物质向籽粒分配。间作玉米净光合速率的提高是通过羧化效率和表观量子效率的提高,促进CO2的固定实现的,而非是光能传递、转化能力的提高。
[1] Liu X H,Han X L,Zhao M Z,Kong Y Z.Studies on solar energy utilization,crop competition and yield analysis in double cropped wheat fields in the North China Plain.Acta Agronomica Sinica,1981,7(1):63-72.
[2] Seran T H,Brintha I.Review on maize based intercropping.Journal of Agronomy,2010,9(3):135-145.
[3] Hugar H Y,Palled Y B.Effect of intercropped vegetables on maize and associated weeds in maize-vegetable intercropping systems.Karnataka Journal of Agricultural Sciences,2008,21(2):159-161.
[4] Li L,Li SM,Sun J H,Zhou L L,Bao X G,Zhang H G,Zhang F S.Diversity enhances agricultural productivity via rhizosphere phosphorus facilitation on phosphorus-deficient soils.Proceedings of the National Academy of Sciences of the United States of America,2007,104(27):11192-11196.
[5] Tsubo M,Walker S,Ogindo H O.A simulation model of cereal-legume intercropping system for semi-arid regions:Ⅱ.Model application.Field Crops Research,2005,93(1):23-33.
[6] Jiao N Y,Ning T Y,Zhao T C,Wang Y,Shi Z Q,Hou L T,Fu GZ,Jiang X D,Li Z J.Characters of photosynthesis in intercropping system of maize and peanut.Acta Agronomica Sinica,2006,32(6):917-923.
[7] Jiao N Y,Zhao T C,Ning T Y,Hou L T,Fu G Z,Li Z J,Chen M C.Effects of maize-peanut intercropping on economic yield and light response of photosynthesis.Chinese Journal of Applied Ecology,2008,19(5):981-985.
[8] Banik P,Sharma RC.Yield and resource utilization efficiency in baby corn-legume-intercropping system in the eastern plateau of India.Journal of Sustainable Agriculture,2009,33(4):379-395.
[9] Huang J Y,Li X P,Sun D L.Ecophysiological effects of multiple cropping of winter wheat-spring corn summer corn in Huanghuaihai Plain.Chinese Journal of Applied Ecology,2003,14(1):51-56.
[10] Zhang J H,Ma Y Y,Wang Z N,Qi J.Research on the improvement of photosynthesis indices of maize in the intercropping system.Journal of Maize Sciences,2006,14(4):104-106.
[11] Arnon D I.Copper enzymes in isolated chloroplasts.Polyphenoloxidase in Beta vulgaris.Plant Physiology,1949,24(1):1-15.
[12] Li X,Feng W,Zeng X C.Advances in chlorophyll fluorescence analysis and its uses.Acta Botanica Boreali-Occidentalia Sinica,2006,26(10):2186-2196.
[13] Wang Z X,Zhu J M,Wang J,Wang Y,Lu Y X,Zheng Q R.The response of photosynthetic characters and biomass allocation of P.bournei young trees to different light regimes.Acta Ecologica Sinica,2012,32(12):3841-3848.
[14] Sun X L,Xu Y F,Ma L Y,Zhou H.A review of acclimation of photosynthetic pigment composition in plant leaves to shade environment.Chinese Journal of Plant Ecology,2010,34(8):989-999.
[15] Gao Y,Duan A W,Liu Z G,Shen X J.Light environment characteristics in maize-soybean strip intercropping system.Chinese Journal of Applied Ecology,2008,19(6):1248-1254.
[16] Gao Y,Duan A W,Liu Z G,Shen X J,Liu ZD,Chen JP.Effect of monoculture and intercropping on radiation use efficiency and yield of maize and soybean.Chinese Journal of Eco-Agriculture,2009,17(1):7-12.
[17] Zhu W X,Zhang H H,Xu N,Wang P,Wang SD,Mu SN,Liang M,Sun GY.Effects of Moru salba and Setaria italica intercropping on their plant growth and diurnal variation of photosynthesis.Chinese Journal of Applied Ecology,2012,23(7):1817-1824.
[18] Du CF,Li CH,Liu T X,Zhao Y L.Response of anatomical structure and photosynthetic characteristics to low light stress in leaves of different maize genotypes.Acta Ecologica Sinica,2011,31(21):6633-6640.
[19] Bi H G,Wang M L,Jiang Z S,Dong X B,Ai X Z.Impacts of suboptimal temperature and low light intensity on the activities and gene expression of photosynthetic enzymes in cucumber seedling leaves.Chinese Journal of Applied Ecology,2011,22(11):2894-2900.
参考文献:
[1] 刘巽浩,韩湘玲,赵明斋,孔杨庄.华北平原地区麦田两熟的光能利用、作物竞争与产量分析.作物学报,1981,7(1):63-72.
[6] 焦念元,宁堂原,赵春,王芸,史忠强,侯连涛,付国占,江晓东,李增嘉.玉米花生间作复合体系光合特性的研究.作物学报,2006,32(6):917-923.
[7] 焦念元,赵春,宁堂原,侯连涛,付国占,李增嘉,陈明灿.玉米-花生间作对作物产量和光合作用光响应的影响.应用生态学报,2008,19(5):981-985.
[9] 黄进勇,李新平,孙敦立.黄淮海平原冬小麦-春玉米-夏玉米复合种植模式生理生态效应研究.应用生态学报,2003,14(1):51-56.
[10] 张建华,马义勇,王振南,齐晶.间作系统中玉米光合作用指标改善的研究.玉米科学,2006,14(4):104-106.
[12] 李晓,冯伟,曾晓春.叶绿素荧光分析技术及应用进展.西北植物学报,2006,26(10):2186-2196.
[13] 王振兴,朱锦懋,王健,汪滢,卢钰茜,郑群瑞.闽楠幼树光合特性及生物量分配对光环境的响应.生态学报,2012,32(12):3841-3848.
[14] 孙小玲,许岳飞,马鲁沂,周禾.植株叶片的光合色素构成对遮阴的响应.植物生态学报,2010,34(8):989-999.
[15] 高阳,段爱旺,刘祖贵,申孝军.玉米和大豆条带间作模式下的光环境特性.应用生态学报,2008,19(6):1248-1254.
[16] 高阳,段爱旺,刘祖贵,申孝军,刘战东,陈金平.单作和间作对玉米和大豆群体辐射利用率及产量的影响.中国生态农业学报,2009,17(1):7-12.
[17] 朱文旭,张会慧,许楠,王鹏,王师丹,牟世南,梁明,孙广玉.间作对桑树和谷子生长和光合日变化的影响.应用生态学报,2012,23(7):1817-1824.
[18] 杜成凤,李潮海,刘天学,赵亚丽.遮荫对两个基因型玉米叶片解剖结构及光合特性的影响.生态学报,2011,31(21):6633-6640.
[19] 毕焕改,王美玲,姜振升,董绪兵,艾希珍.亚适温弱光对黄瓜幼苗光合酶活性和基因表达的影响.应用生态学报,2011,22(11):2894-2900.
猜你喜欢
经济林研究(2022年4期)2023-01-12
广西林业科学(2022年2期)2022-05-09
云南化工(2021年9期)2021-12-21
河北果树(2020年4期)2020-11-26
山东农业科学(2019年11期)2019-12-24
江苏农业科学(2019年11期)2019-07-22
干旱地区农业研究(2017年5期)2017-12-18
新疆农垦科技(2016年2期)2016-08-21
影像科学与光化学(2014年5期)2014-03-11
新疆农垦科技(2014年10期)2014-02-28
