
不同间作模式对山苍子光合特性及其栽培土壤水分和养分的影响
2023-01-12 09:14:46高恩婷曾庆扬谷战英卢雨析冯楠可熊荟璇张春来曾乐景
经济林研究 2022年4期
高恩婷,曾庆扬,谷战英,卢雨析,李 扬,冯楠可,熊荟璇,张春来,曾乐景
(中南林业科技大学 a. 林学院;b. 经济林培育与保护教育部重点实验室;c. 经济林育种与栽培国家林业局重点实验室,湖南 长沙 410004;d. 芦头实验林场,湖南 岳阳 410400)
山苍子Litsea cubeba属于樟科Lauraceae木姜子属Litsea。山苍子作为我国南方地区的特色香料植物资源,具有很高的经济价值。山苍子用途广泛,其根、茎、叶和果实均可入药。湖南省是全国山苍子的主要生产地,该省的山苍子产量约占全国山苍子总产量的94%,湖南省也是山苍子栽培及其遗传多样性的中心[1-3]。发展山苍子产业,对于巩固脱贫成果,实现乡村振兴具有重要意义。
光合作用是植物生长发育的重要影响因素。植物的光合能力可以通过叶片的形态与生理等指标来衡量[4]。林荣呈等[5]指出,在植物的光合产物中,植物干质量的占比为90%~95%。因此,测定植物的光合参数,研究光合参数对植物的影响问题,对于植物的栽培生产具有重要的指导意义。
目前,有关山苍子的研究多集中在山苍子苗木的栽培利用[1]、产业发展[2,6],山苍子油的提取工艺[7]、成分分析[8]、药理学[9]等方面,而关于不同间作模式对山苍子光合作用及其栽培土壤水分和养分的影响的研究报道较少。有关研究结果表明,山苍子林地栽培白车轴草、红车轴草和高羊茅能在一定程度上提高山苍子栽培土壤中水分和养分的含量[10];高群英等[11]的研究结果表明,在干旱胁迫下,百日草和万寿菊作为菊科1年生草本植物,不仅具有观赏性,且其抗旱性也较好,适合作为间作植物,能够对土壤水分产生影响。此外,这几种植物与山苍子间作均可降低除草成本,提高经济效益。为给山苍子间作模式的研究提供理论依据和生产指导,本研究以山苍子同一家系的6年生植株为试材,将其分别与5种植物进行间作试验,测定并分析了不同间作模式对山苍子的光合参数、栽培土壤中水分与养分含量的影响情况,以期从中筛选出能够提高山苍子光合作用效率及土壤水分和养分含量的间作模式。
1 材料与方法
1.1 试验地概况
试验于湖南省株洲市中南林业科技大学山苍子基地进行。试验地属亚热带季风性湿润气候,雨水充沛,光照充足,年无霜期在286 d以上,年平均气温为16~18 ℃,年降水量约为1 500 mm。冬季多西北风,夏季多南风。土壤为红壤,土层厚度为40~80 cm。
1.2 试验设计
试验材料为同一家系的6年生山苍子Litsea cubeba植株;间作植物分别为白车轴草Trifolium repensL.、红车轴草Trifolium pratenseL.、高羊茅Festuca arundinacea、百日草Zinnia elegans和万寿菊Tagetes erectaL.的实生苗,其种子均购于湖南省长沙市红星花卉市场。
试验于2021年3月开始,播种山苍子的间作植物白车轴草、红车轴草、高羊茅、百日草和万寿菊,试验共设6个处理,即清耕(用作对照)、山苍子间作白车轴草、山苍子间作红车轴草、山苍子间作高羊茅、山苍子间作百日草、山苍子间作万寿菊,每个处理各设3个重复。每个处理栽培的山苍子植株各有15株,山苍子的栽培密度均为2.0 m×1.5 m。间作植物的种植方式均为撒播,其播种量均为 75 ㎏·hm-2。
2021年7月中旬,选择晴朗的天气,测定不同间作模式下山苍子叶片的光合参数,同时测定不同间作模式下山苍子栽培土壤中水分与养分的含量。
1.3 测定指标与测定方法
1.3.1 光合参数的测定
使用LI-6400XT便携式光合测定系统测定不同间作模式下山苍子叶片的相关光合参数。每个处理各测定3片叶片,测定的时间段为8:00—18:00时,每隔2 h测定1次,测定指标包括净光合速率(Net photosynthetic rate,Pn)、胞间CO2浓 度(Intercellular CO2concentration,Ci)、气 孔 导 度(Stomatal conductance,Gs)、 蒸 腾速 率(Transpiration rate,Tr)、 气 孔 限 制 值(Stomatal Limitation value,Ls)、光合有效辐射(Photosynthetically active radiation,QPAR)、水分利用效率(Water use efficiency,RWUE)、光能利用效率(Light use efficiency,RLUE)、大气中CO2浓度(Ambient CO2,Ca)等,根据以上指标计算不同间作模式下山苍子的水分利用效率、光能利用效率和气孔限制值[12-15],其计算公式分别如下:
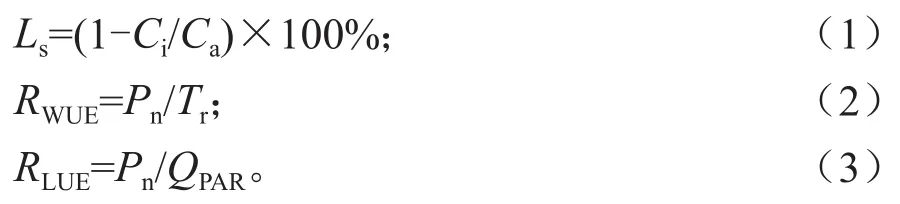
1.3.2 光合-光响应曲线的测定
选择晴天上午的8:30—11:30时,使山苍子叶片在自然光下适应1~2 h,将红蓝光源的光照强度分别设为0、25、50、75、100、150、200、400、600、800、1 000、1 200、1 500 μmol·m-2·s-1,测定不同间作模式下山苍子的光合-光响应曲线,每个处理的测定各设3次重复。以叶子飘的非直角双曲线修正模型[16-18]拟合得到光响应曲线参数。
1.3.3 土壤中水分与养分含量的测定
使用便携式土壤速测仪(型号为PR-ECTHSC-35DC)测定土壤水分含量。
不同间作模式下山苍子栽培土壤样品的采集:按照“S”字形随机选择5个取样点,使用环刀法,挖取土层深度为0~10 cm的土壤样品带回实验室,自然风干后过100目筛后装入自封袋内保存以备用。采用凯氏法测定土壤全氮含量,采用氢氧化钠熔融-钼锑抗比色法测定土壤全磷含量,采用氢氧化钠熔融法测定土壤全钾含量;采用浸提-靛酚蓝比色法测定土壤铵态氮含量,采用盐酸-氟化铵法测定土壤速效磷含量,采用乙酸铵浸提法[19]测定土壤速效钾含量,采用水合热重铬酸钾氧化-比色法[19]测定土壤有机质含量。
1.4 数据处理
采用Excel 2016软件对不同间作模式下山苍子的光合参数、光响应曲线、土壤水分及养分含量等试验数据进行初步分析,采用SPSS 25.0统计软件对山苍子栽培土壤养分含量进行显著性分析,根据叶子飘的非直角双曲线修正模型进行拟合,计算光饱和点、光补偿点等光合参数值,采用Origin8.5软件[20]制图。
2 结果与分析
2.1 环境因子的日变化趋势
山苍子所处环境因子的日变化情况见表1。由表1可知,光照强度,8:00—12:00时持续上升,至 12:00 时达到最大值(1 906.73 μmol·m-2·s-1),而在 18:00 时降至 107.06 μmol·m-2·s-1;大气中的CO2浓度,8:00 时为 406.29 μmol·mol-1,之后逐渐下降,16:00时后逐渐上升;大气温度总体较高,8:00时为37.08 ℃,16:00时达到最高值(39.85 ℃),之后逐渐降低,18:00时降至33.47 ℃;相对湿度,8:00时为43.78%,之后呈下降趋势,14:00时降至最低值(37.15%),随着气温下降,相对湿度略有回升,至18:00时为49.82%。
2.2 不同间作模式下山苍子光合特性的日变化趋势
2.2.1 净光合速率的日变化曲线
不同间作模式下山苍子各光合参数的日变化曲线如图1所示。净光合速率的大小反映了植物光合能力的强弱[21]。由图1a可知,不同间作模式的山苍子净光合速率的日变化曲线均为“双峰型”曲线,说明其均表现出较为明显的“午休”现象。其中,清耕、间作白车轴草、间作高羊茅、间作百日草、间作万寿菊的山苍子的净光合速率均在10:00时出现第1个峰值,其峰值分别为6.62、4.65、7.64、8.64、6.63 μmol·m-2·s-1;随着光强的增强和气温的升高,这5个间作模式的山苍子净光合速率的下降趋势均明显,至14:00时,均下降至较低值;在16:00时又都出现了第2个小峰值,之后山苍子的净光合速率均逐渐下降。但是,间作红车轴草的山苍子净光合速率在12:00时出现了第1个峰值(5.29 μmol·m-2·s-1),而其第 2 个峰值却出现在16:00时。不同间作模式的山苍子净光合速率日均值的大小依次为:间作高羊茅(5.18 μmol·m-2·s-1)>间作百日草(4.89 μmol·m-2·s-1)>间作白车轴草(3.92 μmol·m-2·s-1)>间作万寿菊(3.81 μmol·m-2·s-1)>清 耕(3.78 μmol·m-2·s-1) > 间 作 红 车 轴 草(3.34 μmol·m-2·s-1)。这一观测结果表明,间作高羊茅的山苍子其光合利用能力要强于其他间作模式山苍子的。

表1 环境因子的日变化情况Table 1 Diurnal variation of environmental factors

图1 不同间作模式下山苍子光合参数的日变化曲线Fig. 1 Diurnal variation curves of photosynthetic parameters of L. cubeba with different intercropping patterns
2.2.2 气孔导度的日变化曲线
图1b反映了不同间作模式下山苍子气孔导度的日变化趋势。除了间作百日草的山苍子之外,间作其他植物的山苍子其气孔导度日变化曲线均为“双峰型”曲线,清耕、间作白车轴草、间作高羊茅、间作百日草、间作万寿菊的山苍子其气孔导度的第1个峰值均出现在10:00时,而间作红车轴草的山苍子的第1个峰值却出现在12:00时,比其他间作模式的气孔导度第1个峰值出现的时间延迟2 h。清耕、间作白车轴草、间作红车轴草、间作高羊茅的山苍子其气孔导度第2个峰值均出现于16:00时,而间作万寿菊的山苍子其气孔导度第2个峰值却出现于14:00时。
2.2.3 胞间CO2浓度的日变化曲线
图1c反映了不同间作模式下山苍子的胞间CO2浓度的日变化趋势。6个间作处理的山苍子其胞间CO2浓度均在18:00时达到最大值。清耕、间作白车轴草的山苍子其胞间CO2浓度均在14:00时出现第1个峰值;间作红车轴草的山苍子其胞间CO2浓度分别在10:00和14:00时出现第1个峰值与第2个峰值;间作高羊茅、间作百日草、间作万寿菊的山苍子其胞间CO2浓度均在12:00时出现峰值;16:00时以后,随着环境中光照强度的降低和各处理山苍子净光合速率的降低,不同间作模式的山苍子其胞间CO2浓度均呈逐渐上升趋势,与其净光合速率的日变化趋势均相反。
2.2.4 蒸腾速率的日变化曲线
不同间作模式下山苍子的蒸腾速率日变化趋势如图1d所示。除了间作万寿菊的山苍子之外,其他5个间作模式的山苍子其蒸腾速率日变化曲线均为不对称的“双峰型”曲线。8:00时后,随着气温的逐渐升高与相对湿度的降低,不同间作模式的山苍子其蒸腾速率均呈逐渐增强的变化趋势。清耕、间作白车轴草、间作高羊茅、间作百日草的山苍子其蒸腾速率均在10:00时达到最高值,分别为 1.9、1.1、2.3、2.0 mmol·m-2·s-1;间作红车轴草的山苍子其蒸腾速率却在12:00时才达到峰值(0.6 mmol·m-2·s-1),且其值低于其他 5 个间作处理的。至16:00时,除间作万寿菊的山苍子之外,其他5个间作处理的山苍子其蒸腾速率均出现了第2个峰值;之后,随着光照强度与气温的降低和相对湿度的增加,不同间作模式下山苍子的蒸腾速率均出现了逐渐下降的变化趋势。
2.2.5 气孔限制值的日变化曲线
不同间作模式下山苍子气孔限制值的日变化曲线如图2所示。由图2可知,清耕、间作红车轴草、间作百日草和间作万寿菊的山苍子其气孔限制值日变化曲线均为“双峰型”曲线,12:00时均出现了明显的“午休”现象;间作白车轴草和间作高羊茅的山苍子其气孔限制值日变化曲线均为“单峰型”曲线,间作白车轴草和间作高羊茅的山苍子其气孔限制值均在16:00时达到了最高值,其值分别为64%和77%。

图2 不同间作模式下山苍子的气孔限制值的日变化曲线Fig. 2 Diurnal variation curves of stomatal limitation value (Ls)of L. cubeba with different intercropping patterns
2.2.6 光能利用率和水分利用率的日变化曲线
图3a反映了不同间作模式下山苍子的光能利用效率。由图3a可知,16:00时,不同间作模式的山苍子其光能利用效率均达到了最高值,此时,不同间作模式的山苍子其光能利用效率由大到小依次为:间作白车轴草>间作红车轴草>间作高羊茅>清耕>间作万寿菊>间作百日草。
图3b反映了不同间作模式下山苍子的水分利用效率。由图3b可知,8:00时,间作红车轴草的山苍子其水分利用效率最大,而清耕的最小;之后,随着光照强度的增强、气温的升高和蒸腾速率的增大,不同间作模式下山苍子的水分利用率均呈现下降趋势;14:00—16:00时,除了间作百日草的山苍子外,其他5个间作处理的山苍子其水分利用效率均呈上升趋势,随后均又逐渐下降。至16:00时,不同间作模式的山苍子其水分利用效率由大到小依次为:间作万寿菊>间作高羊茅>间作白车轴草>间作红车轴草>清耕>间作百日草。
2.3 不同间作模式下山苍子的光合-光响应曲线
图4呈现了不同间作模式下山苍子的光响应曲线。由图4可知,当光照强度小于 200 μmol·m-2·s-1时,不同间作模式下山苍子的净光合速率随着光照强度的逐渐增加均呈较快的上升趋势;当光照强度为 200 ~ 400 μmol·m-2·s-1时,除了间作万寿菊的山苍子之外,其他5个间作处理的山苍子其净光合速率的上升趋势均趋平缓;当光照强度大于 400 μmol·m-2·s-1时,清耕、间作白车轴草、间作高羊茅、间作万寿菊的山苍子其光响应曲线均为向下弯曲的曲线;当光照强度大于 800 μmol·m-2·s-1时,间作红车轴草、间作百日草的山苍子其光响应曲线均表现为下降的变化曲线。

图3 不同间作模式下山苍子的光能利用率和水分利用率的日变化曲线Fig. 3 Diurnal variation curves of light energy utilization efficiency and water utilization efficiency of L. cubeba with different intercropping patterns
最大净光合速率反映了叶片的最大光合能力。清耕、间作白车轴草、间作红车轴草、间作高羊茅、间作百日草、间作万寿菊的山苍子最大净光合速率(图4)分别为4.73、4.26、5.79、3.64、7.29、1.91 μmol·m-2·s-1,即间作百日草的山苍子叶片的光合能力最强。

图4 不同间作模式下山苍子的光响应曲线Fig. 4 Light response curves of L. cubeba with different intercropping patterns
根据叶子飘的非直角双曲线修正模型拟合计算出的结果表明:间作红车轴草、间作百日草的山苍子其光饱和点均较高,而其光补偿点均较低,即间作红车轴草、间作百日草均可促使山苍子对光强的适应范围变得更广。
不同间作模式的山苍子其暗呼吸速率由大到小依次为:清耕>间作万寿菊>间作红车轴草>间作白车轴草>间作百日草>间作高羊茅。
不同间作模式的山苍子其表观量子效率由大到小依次为:间作高羊茅>间作红车轴草>间作百日草>间作万寿菊>清耕>间作白车轴草。
2.4 不同间作模式下山苍子栽培土壤的含水量对其光合特性的影响
不同间作模式的山苍子栽培土壤的含水量不同,且山苍子的光合响应特征不同。不同间作模式的山苍子栽培土壤的含水量由大到小依次为:间作万寿菊>间作百日草>间作高羊茅>间作红车轴草>清耕>间作白车轴草。其中,间作白车轴草的土壤含水量最低,为13.38%;间作万寿菊的土壤含水量最高,为16.60%。
不同间作模式的山苍子栽培土壤的含水量对其光合特性的影响情况如图5所示。由图5a可知,当光照强度小于 200 μmol·m-2·s-1时,不同间作模式的山苍子其蒸腾速率均呈较快上升的变化趋势,其中,间作百日草的山苍子其蒸腾速率的上升趋势最快;间作万寿菊的山苍子的蒸腾速率在光照强度大于 400 μmol·m-2·s-1时呈缓慢下降趋势,而在光照强度大于 600 μmol·m-2·s-1时却趋于平缓。随着光照强度的增加,不同间作模式的山苍子其蒸腾速率(图5a)和气孔导度(图5d)均呈上升趋势,其中,土壤含水量最高的间作万寿菊的山苍子其气孔导度和蒸腾速率的上升趋势均最缓慢,而土壤含水量较高的间作高羊茅的山苍子其气孔导度和蒸腾速率均呈平稳变化趋势。当光照强度小于 200 μmol·m-2·s-1时,不同间作模式的山苍子其胞间CO2浓度(图5b)均呈下降趋势,而其水分利用效率(图5c)均呈上升趋势;当光照强度大于 400 μmol·m-2·s-1时,其胞间 CO2浓度均呈缓慢上升的变化趋势,而其水分利用效率均呈缓慢下降趋势,其中土壤含水量最大的间作万寿菊的山苍子其胞间CO2浓度的上升速度要快于其他间作模式的。
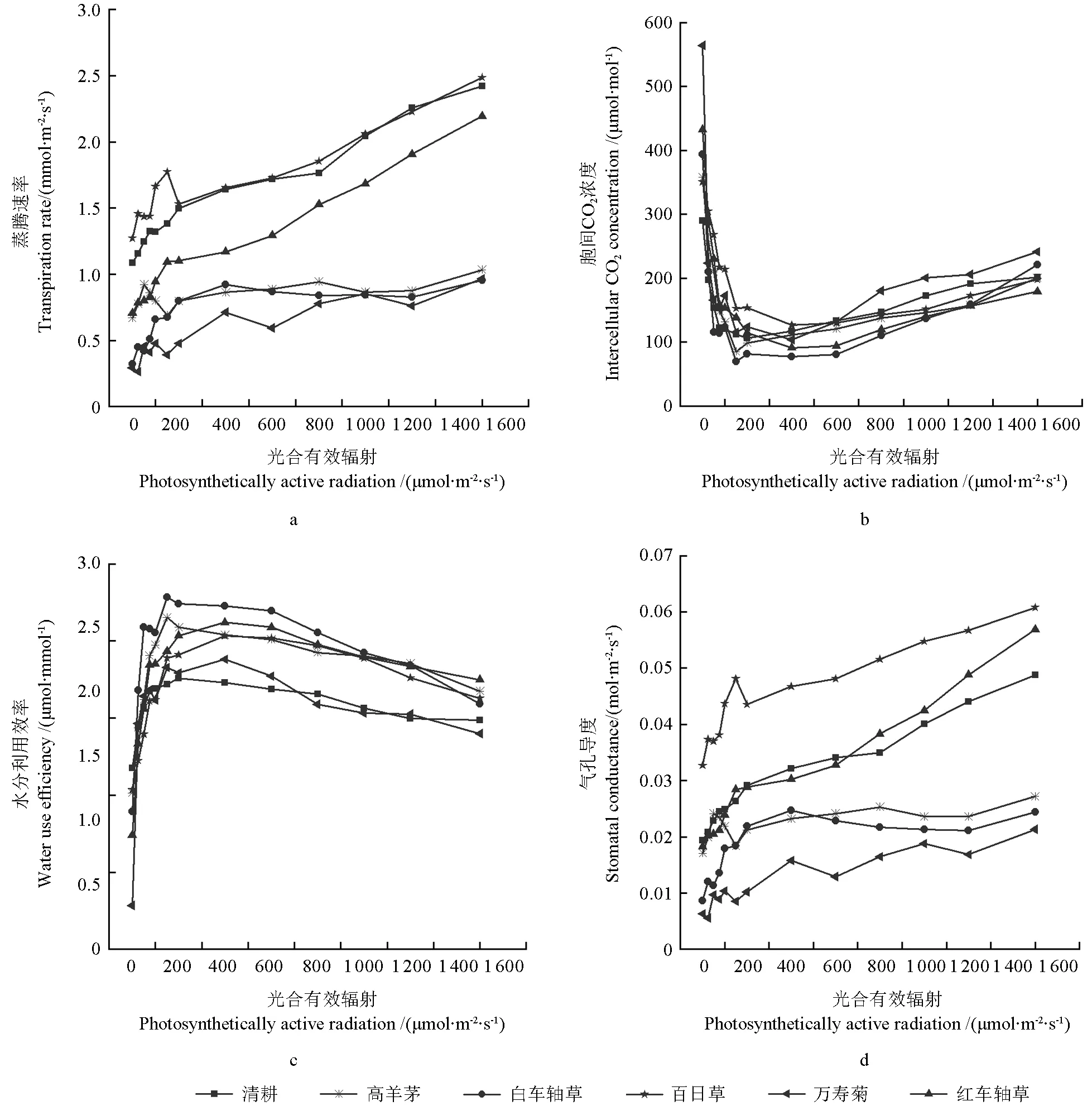
图5 不同间作模式下山苍子栽培土壤的含水量对其光合特性的影响Fig. 5 Effects of soil water content on photosynthetic characteristics of L. cubeba under different intercropping patterns
2.5 不同间作模式下山苍子栽培土壤的养分含量
不同间作模式的山苍子栽培土壤0~10 cm土层中的养分含量见表2。表2表明,间作万寿菊、清耕与其他间作处理之间土壤中的全氮含量均有显著差异,不同间作模式的山苍子栽培土壤中的全氮含量由大到小依次为:间作百日草>间作高羊茅>间作万寿菊>间作白车轴草>间作红车轴草>清耕。清耕土壤中的全磷含量,除了与间作白车轴草的差异不显著之外,而与其他各间作处理之间土壤全磷含量均有显著性差异;间作白车轴草的土壤全磷含量较清耕的高出20.83%。清耕土壤中的全钾含量最高,且其与间作白车轴草和间作红车轴草的土壤全钾含量间的差异均不显著。清耕与其他各间作处理之间土壤中有机质的含量差异均显著;而间作白车轴草和间作红车轴草之间土壤有机质含量的差异性不显著;其余3种间作模式之间土壤有机质的含量差异均显著,其中间作红车轴草的土壤有机质含量较清耕的高出62.79%。清耕与间作白车轴草、间作万寿菊之间土壤铵态氮的含量差异均不显著;间作红车轴草和间作高羊茅的土壤铵态氮含量,与清耕的相比,分别高出27.58%与25.39%。清耕与间作高羊茅、间作万寿菊之间土壤速效磷的含量差异均不显著;其他间作模式的土壤速效磷含量均高于清耕的,其中间作白车轴草和间作百日草的土壤速效磷含量,与清耕的相比,分别高出72.02%与39.01%。清耕和间作白车轴草之间土壤速效钾的含量差异不显著,不同间作模式下山苍子栽培土壤中的速效钾含量从大到小依次为:间作百日草>间作高羊茅>间作万寿菊>间作红车轴草>间作白车轴草>清耕。
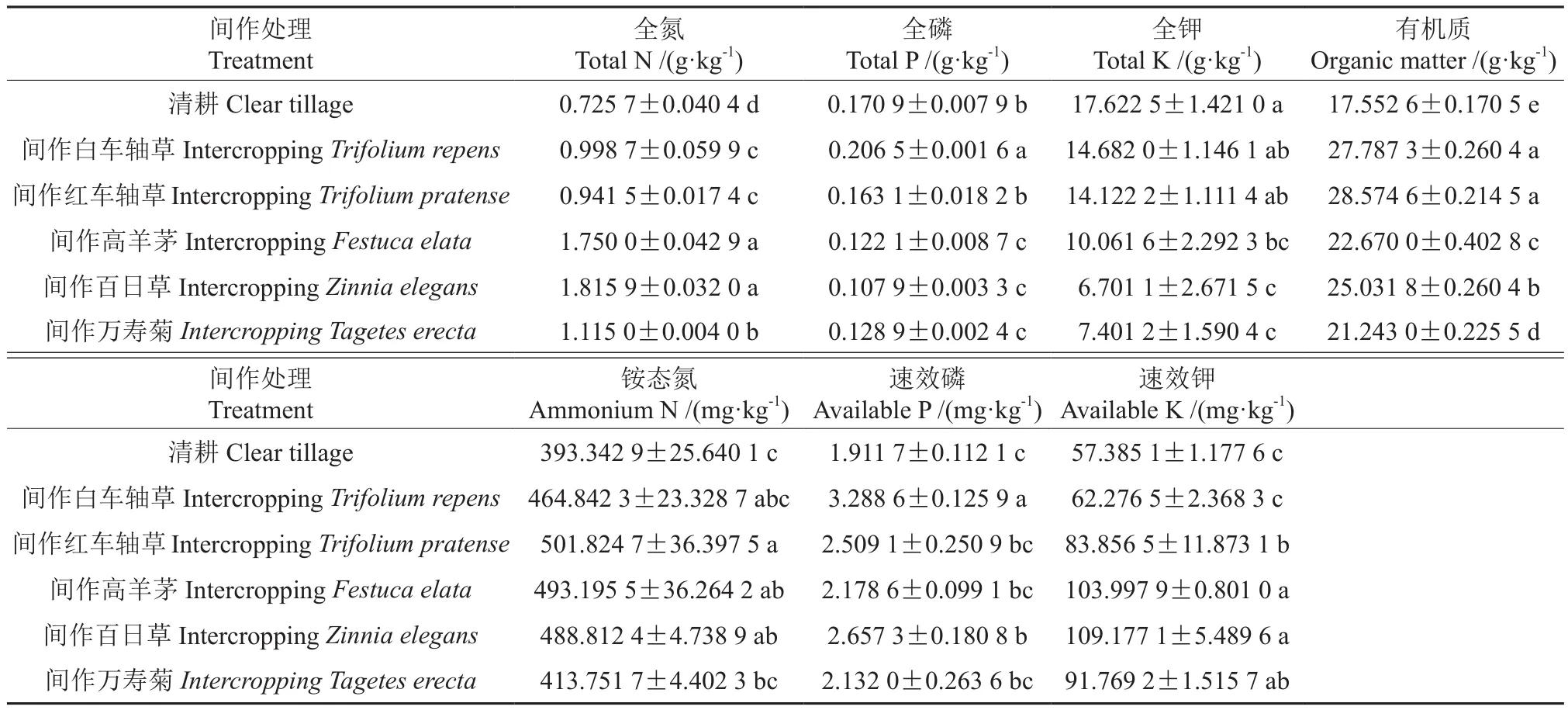
表2 不同间作模式的山苍子栽培土壤0~10 cm土层中的养分含量†Table 2 Soil nutrient of L. cubeba with different intercropping patterns
3 讨 论
3.1 不同间作模式下山苍子净光合速率的日变化曲线
植物的光合速率既受植物本身控制,也受外界环境的影响[12]。研究结果表明,清耕、间作白车轴草、间作红车轴草、间作高羊茅的山苍子其净光合速率均在16:00时才达到最大值,而间作百日草、间作万寿菊的山苍子其净光合速率均在10:00时就早已达到最大值,比较而言,16:00时的最大值明显高于10:00时的最大值,且其净光合速率的变化曲线均为“双峰型”曲线。“午休”现象是植物对外界环境作出的生理反应[21]。研究中发现,清耕、间作白车轴草、间作红车轴草、间作高羊茅的山苍子均在14:00时才出现明显的“午休”现象,而间作百日草和万寿菊的山苍子均在12:00时便出现了明显的“午休”现象。与其他间作处理相比,间作高羊茅、间作百日草均可延缓山苍子“午休”现象的出现。随着气温的升高,不同间作模式的山苍子其净光合速率并非直线上升,且各个间作处理之间山苍子的净光合速率存在差异,间作红车轴草的山苍子其净光合速率峰值最大。
有关研究结果表明,光合作用受到气孔因素和非气孔因素的限制。在气孔导度和胞间CO2浓度均下降时,此时的气孔限制引起了净光合速率的下降,即为气孔因素限制了光合作用;反之,若胞间CO2浓度升高,净光合速率下降,即为非气孔因素限制了光合作用[17,22]。分析不同间作模式下山苍子的净光合速率、气孔导度、胞间CO2浓度日变化曲线可知,山苍子的净光合速率随着气孔导度的下降和胞间CO2浓度的升高而降低,说明不同间作模式下山苍子的光合作用均受到了非气孔因素的限制。因此,在山苍子的栽培管理中,间作高羊茅、间作百日草均可有效增强山苍子的光合能力,延缓其“午休”现象的出现,这两种间作模式均有利于山苍子植株的生长发育。
3.2 不同间作模式下山苍子蒸腾速率、气孔导度、胞间CO2浓度、气孔限制值的日变化曲线
清耕、间作白车轴草、间作红车轴草、间作高羊茅的山苍子其蒸腾速率和气孔导度日变化曲线都表现为显著的“双峰型”曲线,而间作百日草和间作万寿菊的山苍子其蒸腾速率和气孔导度日变化曲线却表现为“单峰型”曲线或次峰不明显的变化曲线;其中,间作高羊茅和间作红车轴草的山苍子其蒸腾速率峰值出现在12:00时,说明间作高羊茅、间作红车轴草均可使山苍子在高温和高光强下仍能保持较高的蒸腾速率,使其耗散过多的热量,有效降低山苍子的叶温。
气孔导度表示叶片气孔张开的程度,是反映叶片传导能力的关键指标,其传导能力在一定程度上影响着植物的光合速率、蒸腾速率和呼吸速率[21,23]。8:00—10:00时,不同间作模式下山苍子的气孔导度随着气温和光照强度的上升均呈逐渐增大的变化趋势,其中,间作白车轴草、间作百日草的山苍子其气孔导度的增大趋势均较平缓,这一结果说明,与其他几个间作处理的山苍子叶片相比,间作白车轴草、间作百日草的山苍子叶片对气温和光强均不敏感。
清耕、间作白车轴草、间作红车轴草、间作高羊茅的山苍子其胞间CO2浓度在16:00时均呈先下降后上升的变化趋势,其主要原因是,16:00时的气温较高而相对湿度较低,山苍子为减小蒸腾作用关闭叶片气孔,使其胞间CO2浓度降低。而间作百日草和间作高羊茅的山苍子其胞间CO2浓度的变化均较平缓。有关研究结果表明,植物在中午时其净光合速率下降的主要原因有气孔限制和非气孔限制两种。当胞间CO2浓度下降、气孔限制值增加时,此时植物的净光合速率下降,说明是由气孔限制引起的[11,22]。因此,间作百日草的山苍子其净光合速率在12:00—14:00时的变化曲线、间作白车轴草的山苍子其净光合速率在8:00—10:00时的变化曲线、间作高羊茅的山苍子其净光合速率分别在8:00—10:00与12:00—14:00时的变化曲线、间作百日草的山苍子其净光合速率在14:00—16:00时的变化曲线,都是因为受到山苍子的气孔限制而形成的;清耕、间作红车轴草、间作万寿菊的山苍子在8:00时后其净光合速率和气孔导度均呈下降趋势,而其胞间CO2浓度却呈上升趋势,说明此时山苍子叶片利用二氧化碳的能力有所下降,这可能因为光照强度的增加促使叶片温度升高和植物细胞气孔扩张,使得叶片内相关酶活性降低和细胞间物质交换加快,叶片利用二氧化碳的能力因而减弱了[23-24]。因此,引起清耕、间作红车轴草、间作万寿菊的山苍子净光合速率下降的原因主要为非气孔限制。
3.3 不同间作模式下山苍子的光合-光响应曲线
本研究采用非直角双线修正模型拟合得到的光合-光响应曲线(图4)表明,间作白车轴草和间作百日草的山苍子其最大净光合速率与光饱和点均最高,即间作白车轴草、间作百日草均可增强山苍子对强光的适应性。植物对弱光的利用能力可以通过表观量子效率和光补偿点来体现[21]。研究结果还表明,山苍子的表观量子效率,间作高羊茅和间作红车轴草的均高于其他间作处理的;而山苍子的光补偿点,间作高羊茅和间作红车轴草的均低于其他间作处理的。这一结果说明,间作高羊茅、间作红车轴草均可提高山苍子利用弱光的能力。综上所述,间作高羊茅和间作红车轴草的山苍子的光利用范围比其他间作处理山苍子的都要宽,说明间作高羊茅和间作红车轴草均可促使山苍子能较好地适应强光和弱光环境。
3.4 不同间作模式下山苍子的光能利用效率和水分利用效率
光能利用效率的大小可以反映植物通过进行光合作用转化有机物效率的大小[12]。各间作处理山苍子的光能利用效率在16:00时均达到了各自的峰值,其中,间作白车轴草、间作红车轴草、间作高羊茅的山苍子的光能利用效率均高于清耕、间作百日草和间作万寿菊的山苍子的,说明间作红车轴草、间作白车轴草、间作高羊茅均可促使山苍子能较好地适应气温低、光照强度低的环境。
水分利用效率的大小可以体现植物抗旱能力的强弱[21]。在一天之内温度最高的16:00时,间作高羊茅和间作万寿菊的山苍子其水分利用效率均最高,说明其用于光合作用的有效水分都较多,其抗旱能力均较强于其他间作处理山苍子的。
3.5 不同间作模式的山苍子栽培土壤含水量与养分含量的差异分析
研究结果表明,不同间作模式的山苍子栽培土壤的含水量不同。当光照强度小于200 μmol·m-2·s-1时,随着光照强度的增加,土壤含水量高的间作模式的山苍子其气孔导度和蒸腾速率的上升趋势均较缓慢,其中,间作万寿菊的山苍子其气孔导度和蒸腾速率的上升趋势均最缓慢;而当光照强度大于400 μmol·m-2·s-1时,土壤含水量越大,其对应间作模式的山苍子胞间CO2浓度的上升速度则越快;当光照强度大于 600 μmol·m-2·s-1时,不同间作模式的山苍子的净光合速率均趋于稳定,其胞间CO2浓度、蒸腾速率和气孔导度均有不同程度的上升,土壤含水量越低的间作模式,其上升速度越快,说明较大的土壤含水量可以更有效地降低山苍子叶片的温度,并能调节山苍子光合器官的活性,使其能够更好地适应较强的光照强度[22]。
研究结果还表明,不同间作模式的山苍子栽培土壤0~10 cm土层中的养分含量存在较为显著的差异。其中,间作万寿菊的山苍子栽培土壤中的全氮含量最高,且其含量与清耕的相比有显著性差异,而清耕的山苍子栽培土壤中的全氮含量最低,说明间作百日草、间作高羊茅、间作万寿菊、间作白车轴草和间作红车轴草均能不同程度地提高山苍子栽培土壤中的全氮含量;间作白车轴草的山苍子栽培土壤中的全磷含量最高,而其他间作模式的土壤全磷含量均低于清耕的;不同间作模式均未能显著提高土壤中的全钾含量;间作红车轴草、间作白车轴草、间作百日草、间作高羊茅和间作万寿菊均能不同程度地提高山苍子栽培土壤中的有机质含量;间作红车轴草和间作高羊茅均能显著提高山苍子栽培土壤中的铵态氮含量;间作白车轴草和间作百日草均能较为显著地提高山苍子栽培土壤中的速效磷含量;间作百日草、间作高羊茅、间作万寿菊和间作红车轴草均能显著地提高山苍子栽培土壤中的速效钾含量。
4 结 论
不同间作模式对山苍子的光合特性、土壤水分和养分均有不同程度的影响,选择适宜的间作模式可以有效提高山苍子的光合能力及其栽培土壤中水分和养分的含量,使山苍子能够更好地适应强光环境,并可为其提供生长所需的土壤水分和养分。本研究的局限性在于只分析了不同间作模式对山苍子光合特性及其栽培土壤水分和养分的影响情况,而不同间作模式下山苍子的生长发育、果实品质及精油含量等指标是否发生改变,这一问题有待进一步研究。此外,本研究仅在湖南省株洲市中南林业科技大学的山苍子基地内进行,至于采用相同间作模式栽培的山苍子在其他地区的光合作用及其栽培土壤中水分和养分含量等情况是否与此基地内的一致,也尚待试验研究。
猜你喜欢
河北农业(2023年10期)2023-11-28 09:23:34
食品工业(2022年12期)2023-01-12 07:18:32
湖北农业科学(2022年23期)2022-02-13 14:53:02
云南农业(2021年9期)2021-09-24 11:56:56
湖南农业大学学报(自然科学版)(2021年2期)2021-05-06 00:21:38
农业与技术(2020年2期)2020-02-05 07:08:52
中国粮油学报(2019年4期)2019-07-12 09:06:40
山西农经(2019年5期)2019-02-22 09:20:54
中国新技术新产品(2016年19期)2016-12-12 10:21:42
现代农业(2016年5期)2016-02-28 18:42:34
