
水貂犬瘟热病毒受体SLAM的原核表达
2013-08-14 08:01张洪亮靳继惠王怡男
动物医学进展 2013年2期
王 聪,张洪亮,黄 娟,靳继惠,王怡男,单 虎
(青岛农业大学山东省预防兽医学重点实验室,青岛266109)
犬瘟热(Canine distemper,CD)是由犬瘟热病毒(Canine distemper virus,CDV)引起犬科(犬、狐、貉等)、鼬科(水貂、雪貂、黄鼬等)、灵猫科(果子狸)及一部分浣熊科动物的一种急性、亚急性、接触性传染病,是当前对养犬业、毛皮动物养殖业和野生动物保护业危害最大的疫病之一[1]。研究表明,CDV具有比以前设想的更广的宿主范围[2],免疫动物暴发犬瘟热的病例在世界多个国家和地区也都有报道[3]。细胞表面受体为病毒的组织嗜性和感染宿主范围的决定因素,CDV的跨种间传播现象与其感染宿主的细胞受体的表达有着密切关系[2]。因此,研究CDV受体的特性、功能及其与病毒蛋白相互作用的机制,不仅有助于阐述病毒致病机理,对研制更有效的抗病毒药物、疫苗和诊断试剂也具有重大的理论与现实意义。
Tatsuo H等[4]发现人的信号淋巴细胞激活因子(Signal lymphocyte activation molecule,SLA M),又称CD150,为麻疹病毒感染的细胞受体后,其又通过CDV野毒株在稳定表达SLAM的CHO细胞上感染试验相继证实犬的SLAM为CDV感染胞上高水平表达,对调控机体免疫系统抗体和各种免疫因子产生具有重要作用[7]。因此,CDV的感染能造成动物机体免疫系统的破坏,从而致使动物因免疫缺陷而感染其他病原造成较高的病死率。受体SLAM的分子差异决定了麻疹病毒属病毒的宿主嗜性[8]。但是,目前关于CDV与受体SLAM 相互作用的具体机制还不完全清楚。
本研究通过将水貂的SLAM基因连接到高效表达载体pGEX-6p-1,将原核重组质粒于E.coli BL21宿主菌中进行诱导表达,并对其进行定性鉴定。
1 材料与方法
1.1 材料
克隆质粒pMD18-T-SLAM(含水貂SLAM基因)由中国农业科学院特产研究所馈赠;pGEX-6p-1原核表达质粒,感受态大肠埃希菌(E.coli)DH5α、BL21均为笔者所在实验室保存;限制性内切酶EcoRⅠ、SalⅠ,SolutionⅠ均购自宝生物工程(大连)有限公司;San Prep柱式DNA胶回收试剂盒购自上海生工生物工程技术服务有限公司,质粒小剂量提取试剂盒购自美国Omega Bio-Tek公司;鼠抗GST标签单克隆抗体、兔抗鼠IgG(HRP)购自上海酶联生物科技有限公司;一站式GST标记蛋白质微量纯化套装购自北京天恩泽基因科技有限公司。
1.2 方法
1.2.1 引物设计与目的基因扩增 根据GenBank收录的SLAM序列(登录号:FJ626692),利用Primer 5.0软件设计一对特异性引物,并在引物上、下游的5′端分别引入EcoRⅠ和SalⅠ酶切位点,引物序列 为:上 游 引 物:5′-CGGAATTCATGGATCCCAAGAGGCTTCT-3′, 下 游 引 物:5′-GC GTCGACTCAGCTCTCTGGAAGTGTCAC-3′(下划线处为EcoRⅠ和SalⅠ酶切位点)。PCR产物大小预计约为1 035bp。
以质粒pMD18-T-SLAM为模板,进行PCR反应。PCR扩增体系为25μL,其中10×PCR buffer 2.5μL,上、下游引物各0.5μL,dNTP Mixture 2μL,模板2μL,ddH2O 17μL,TaqDNA 聚合酶0.5μL。PCR反应条件为:95℃预变性3min;94℃30s,55℃1min,72℃1min,35个循环;72℃延伸10min。
1.2.2 重组原核表达质粒pGEX-6p-1-SLAM的构建
将PCR产物用10g/L琼脂糖凝胶电泳回收,进一步用San Prep柱式DNA胶回收试剂盒纯化DNA。采用限制性内切酶EcoRⅠ和SalⅠ对PCR产物和pGEX-6p-1分别进行双酶切,酶切产物用SolutionⅠ于16℃连接过夜,连接产物转化DH5α感受态细胞。重组表达载体的鉴定采用PCR及双酶切鉴定,并送北京六合华大基因科技(青岛)股份有限公司测序。
1.2.3 SLAM-GST在大肠埃希菌BL21中的表达
将重组质粒pGEX-6p-1-SLAM转化感受态细胞BL21,挑取单个克隆接种于LB液体培养基,37℃、220r/min振荡过夜。取过夜的菌液按1∶100比例接种于新鲜的LB液体培养基,37℃剧烈振荡2h~3h,待菌液浓度达OD 600nm=0.6~0.8时,加入异丙 基硫 代-β-D-半 乳 糖 苷 (IPTG)至 终 浓 度 为1.0mmol/L,于 37℃ 诱 导 5h,取 样 进 行 SDSPAGE电泳。同时设BL21/pGEX-6p-1对照及未诱导的BL21/pGEX-6p-1-SLAM对照。
1.2.4 表达产物的纯化 将诱导表达后的BL21/pGEX-6p-1-SLAM菌液经GST标记蛋白质微量纯化套装进行纯化,分别取纯化前和纯化后产物进行SDS-PAGE电泳。
1.2.5 表达产物的 Western blot分析 分别诱导BL21/pGEX-6p-1-SLAM 和 BL21/pGEX-6p-1,取诱导后的菌液及未诱导的菌液进行SDS-PAGE电泳,采用电转印法转印至PVDF膜,220V,2h,取出转印膜,用100g/L脱脂奶粉封闭过夜,加入1∶1 000鼠抗GST标签单克隆抗体,孵育4h,再加入1∶2 000稀释的羊抗鼠-HRP二抗,DAB显色。
2 结果
2.1 目的基因的扩增
水貂SLAM基因的PCR扩增产物在10g/L琼脂糖凝胶中可见到一条1 035bp左右的特异性条带(图1)。

图1 PCR扩增SLAM基因Fig.1 SLAM gene amplified by PCR
2.2 重组原核表达质粒pGEX-6p-1-SLAM的构建与鉴定
将构建的重组质粒pGEX-6p-1-SLAM用EcoRⅠ和SalⅠ双酶切,经10g/L琼脂糖凝胶电泳出现2条特异条带,其中一条大小为4 984bp,另一条为1 035bp左右。表明目的基因已经插入到质粒pGEX-6p-1的相应克隆位点中(图2),DNA序列分析结果表明,插入片段具有正确的阅读框。
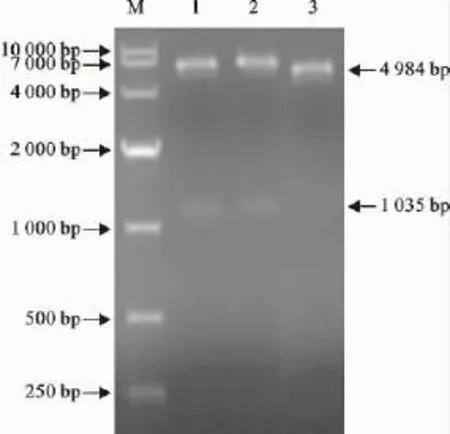
图2 重组质粒pGEX-6p-1-SLAM双酶切鉴定Fig.2 The recombinant plasmid pGEX-6p-1-SLAM digested by restriction enzymes
2.3 SLAM-GST在大肠埃希菌中的表达
pGEX-6p-1-SLAM重组转化菌经IPTG于37℃诱导5h后,细菌裂解上清液在66.4ku附近出现一蛋白条带,与预期大小(63.2ku)基本一致,而空载体转化菌经诱导可表达26ku的GST蛋白(图3)。
2.4 SLAM-GST的纯化
经纯化获得较纯的重组蛋白(图4)。

图3 SDS-PAGE鉴定目的蛋白的表达Fig.3 The expressed protein identified by SDS-PAGE
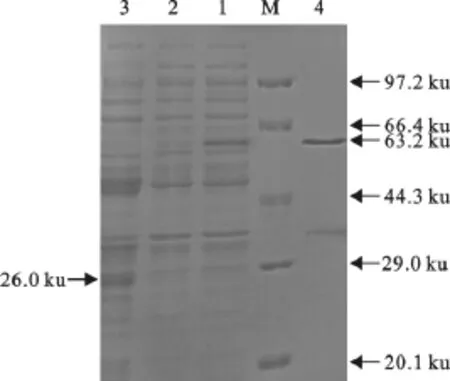
图4 重组蛋白的纯化Fig.4 Purification of recombinant protein
2.5 Western blot分析
结果显示,SLAM-GST融合蛋白能与鼠抗GST标签单克隆抗体发生特异性反应(图5),表明该表达蛋白具有良好的抗原性。
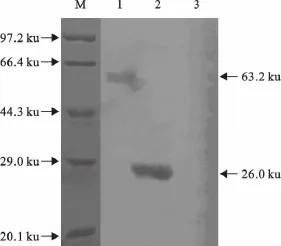
图5 Western blot鉴定目的蛋白的表达Fig.5 The expressed protein identified by Western blot
3 讨论
SLAM作为犬瘟热病毒受体的鉴定有助于我们更好地理解该病毒感染的致病机理,特别是犬瘟热病毒介导的免疫缺陷机理。SLAM受体是表达在淋巴细胞上的细胞膜糖蛋白,犬瘟热病毒感染水貂后能与其特异性结合,支持犬瘟热病毒复制,能引起细胞病变效应[7]。SLAM结构中的V结构域为CDV H蛋白识别并与之结合所必需的,其在传递来自抗原递呈细胞提供的信息及调控免疫细胞分泌抗体和细胞因子方面起到重要作用[9],但是,该结构域中发挥受体功能的关键氨基酸残基位点还需进一步研究。本研究根据GenBank收录的SLAM基因序列设计了一对特异性引物,利用RT-PCR克隆到水貂SLAM基因编码区,选用原核表达载体进行表达,该蛋白的获得为SLAM受体与犬瘟热病毒结构蛋白的相互作用研究奠定了基础,从而为设计、开发病毒受体阻断剂提供了理论依据。
pGEX-6p-1是一个高效表达载体,且该载体表达的GST融合蛋白可特异性结合GSH,为用GSTrap FF亲和层析柱纯化表达产物提供了方便[10]。因此,选用这一载体来表达融合蛋白。含重组质粒的大肠埃希菌经IPTG诱导,其菌体蛋白经SDS-PAGE电泳,在分子质量63.2ku处出现大量蛋白,这与推算的融合蛋白的大小一致。用Western blot检测GST融合蛋白,结果显示该融合蛋白具有良好的抗原性。在今后的研究中,可进一步应用制备的SLAM蛋白免疫动物获得高免血清,并采用免疫组化法对SLAM受体在水貂不同组织分布情况及与病毒载量的关系等进行深入研究。
[1] Kameo Y,Nagao Y,Nishio Y,et al.Epizootic canine distemper virus infection among wild mammals[J].Vet Microbiol,2012,154(3-4):222-229.
[2] Goodrich J M,Quigley K S,Lewis J C,et al.Serosurvey of free-ranging Amur tigers in the Russian Far East[J].J Wildl Dis,2012,48(1):186-189.
[3] McCarthy A J,Shaw M A,Goodman S J.Pathogen evolution and disease emergence in carnivores[J].Proc Biol Sci,2007,274(1629):3165-3174.
[4] Tatsuo H,Ono N,Tanka K,et al.SLAM(CDwl50)is a cellular receptor for measles virus[J].Nature,2000,406:893-897.
[5] Tatsuo H,Ono N,Yanagi Y.Morbilliviruses use signaling lymphocyte activation molecules(CDl50)as cellular receptors[J].J Virol,2001,75:5842-5850.
[6] Tatsuo H,Yanagi Y.The morbillivirus receptor SLAM (CDl50)[J].Microbiol Immunol,2002,46(3):135-142.
[7] Ostrakhovitch E A,Li S S.The role of SLAM family receptors in immune cell signaling[J].Biochem Cell Biol,2006,84(6):832-843.
[8] 赵建军,张海玲,高 晗,等.狐、貉和水貂犬瘟热病毒受体SLAM的基因克隆及其真核表达[J].兽类学报,2010,30(1):79-86.
[9] 刘 鑫.麻疹病毒通过受体SLAM对昆虫细胞的感染及其诱发细胞凋亡的机制研究[D].湖北武汉:武汉大学,2005.
[10] 顾小雪,李玉峰,姜 平,等.猪生殖与呼吸综合征病毒GP5蛋白抗原表位基因的表达及其表达产物的抗原性[J].中国兽医科学,2007,37(12):1053-1057.
猜你喜欢
江西农业学报(2021年4期)2021-04-20
环球时报(2020-06-08)2020-06-08
特产研究(2016年3期)2016-04-12
特产研究(2016年3期)2016-04-12
西南医科大学学报(2015年1期)2015-08-22
中国当代医药(2015年9期)2015-03-01
西南军医(2015年6期)2015-01-23
当代畜禽养殖业(2014年5期)2014-08-31
养殖与饲料(2014年10期)2014-02-28
云南畜牧兽医(2014年2期)2014-02-28
