
二疣犀甲对不同寄主茎干的产卵选择
2013-04-04 03:19钟宝珠吕朝军李洪王东明覃伟权
环境昆虫学报 2013年1期
钟宝珠,吕朝军*,李洪,王东明,覃伟权,王 智
(1.中国热带农业科学院椰子研究所,海南 文昌 571339[2.海南省森林资源监测中心,海口 570203)
二疣犀甲Oryctes rhinoceros(Linnaeus)又名椰蛀犀金龟,隶属于鞘翅目Coleoptera、犀金龟科Dynastidae,是亚洲及南太平洋椰子种植区危害椰子树的一种重要害虫。二疣犀甲寄主植物种类丰富,包括椰子Cocos nucifera、油棕Elaeis guineensis、桄榔Arenga pinnata、大王棕 Roystonea regia、甘蔗Saccharum officinarum等多种植物。其中椰子、油棕是最主要的寄主植物 (Dhileepan,1988)。
对昆虫产卵活动进行研究,能反映昆虫与植物相互作用的某些特点,同时可阐明植食性昆虫对植物的利用策略,从而影响昆虫种群的繁衍(钦俊德,1987)。据文献报道,在马来西亚二疣犀甲取食椰子和其他31种棕榈植物 (Cumber,1957),大量死亡油棕茎干的堆积造成的腐殖质,成为二疣犀甲的理想繁衍场所 (Mohd Hashim,1993),这些腐殖质为后期二疣犀甲种群的迅速扩大提供了基础。在中国海南省调查发现,死亡的油棕和椰子木段上均可采集到大量的二疣犀甲幼虫,同时在掺杂一定量幼虫粪便的已取食木段上仍可看到初孵化的幼虫。近年来,利用调节昆虫取食、交配、产卵等行为的信息化学物质来发展新的防治技术越来越受到广泛重视 (孙江华等,2000;张建平等,2000)。目前关于二疣犀甲在不同寄主腐殖质之间是否存在偏好,以及不同原因造成的寄主死亡堆积物,在诱集二疣犀甲产卵方面是否存在差异等方面是否研究尚属空白,如能在此方面进行深入探讨,将有利于明确二疣犀甲的产卵选择偏好性,为二疣犀甲的防控提供参考。
1 材料与方法
1.1 供试材料
虫源:二疣犀甲成虫采集于田间,转移至室内后以雌雄比为2∶1配对后置于生化培养箱中(26℃,RH 75% ~95%,10L∶14D)以成熟香蕉切片饲养。具体饲养方法参考Bedford(1976)所述。15 d后,挑选健康个体供实验。
产卵箱:尺寸为60 cm×50 cm×50 cm。根据处理数量将箱底平均分配后进行间隔,然后铺上不同产卵基质,基质厚度为15 cm。基质上覆盖一层薄木片供二疣犀甲爬行和交配,薄木片上放置纵向切开的成熟香蕉供二疣犀甲自由取食。
孵化箱:尺寸为25 cm×15 cm×10 cm。底部铺6 cm厚的相应产卵基质供二疣犀甲的卵孵化。
1.2 试验方法
1.2.1 对不同腐烂阶段椰子茎干的选择
以海南高种椰子为供试寄主,田间采集正常茎干、半腐烂茎干和充分腐烂的茎干,粉碎后作为产卵基质。每个养虫笼中接入二疣犀甲雌成虫12头、雄成虫6头,任其自由选择产卵基质,并饲以成熟香蕉切片。每处理重复5次,隔5 d统计一次产卵箱中卵的数量,连续统计25 d。将所产虫卵用湿润毛笔小心从产卵箱中分离后,迅速转移至放有相应产卵基质的孵化箱 (25 cm×15 cm×10 cm)中进行孵化,统计卵孵化率。
1.2.2 对5种棕榈科植物的产卵选择
选择海南高种椰子、油棕、槟榔、大王棕、蒲葵等5种处于半腐烂状态的棕榈植物茎干,粉碎后分别放入产卵箱中作为产卵基质。实验处理方法同1.2.1。
1.2.3 对不同死因椰子茎干的产卵选择
选择因感染椰子茎泻血病或茎干腐烂病致死、虫害致死和台风折断致死,且呈半腐烂状态的海南高种椰子树干粉碎物为产卵基质。实验处理方法同1.2.1。
1.2.4 对5种椰子品种的产卵选择
选择台风等自然伤害致死的、半腐烂状态的椰子茎干粉碎物为基质进行试验,椰子品种包括海南高种椰子、黄矮椰子 (Yellow dwarf)、红矮椰子 (Red dwarf)、香水椰子 (Aromatic green dwarf)和马哇椰子 (Mawa)茎。实验处理方法同1.2.1。
1.2.5 幼虫粪便对成虫产卵的影响
选择半腐烂状态的海南高种椰子树干粉碎物作为产卵基质,分别在基质中加入不同比例的二疣犀甲幼虫粪便,使得产卵基质中幼虫粪便最终比例分别为0、10%、25%、40%、55%(主要是为了测试茎干内已有幼虫危害,且茎干被吃掉一半的状态下成虫是否会选择产卵),并将幼虫粪便与茎干粉碎物混匀后用于试验。实验处理方法同1.2.1。
1.3 数据处理
本文结果均采用平均值±标准误 (M±SE)来表示,并采用SPSS13.0软件进行差异显著性分析。
2 结果与分析
2.1 对不同腐烂阶段椰子茎干的选择
二疣犀甲对不同腐烂阶段的海南高种椰子茎干的产卵选择结果如表1所示。结果表明,二疣犀甲对不同腐烂状态椰子茎干的产卵选择性具有显著差异,其中以半腐烂状态的茎干对二疣犀甲的产卵引诱效果最强,单雌平均产卵数为27.06粒,其次为完全腐烂的椰子茎干,而正常椰子茎干粉碎物引诱效果最差。
不同腐烂状态的椰子茎干对二疣犀甲卵的孵化率亦有影响。其中以半腐烂状态的椰子茎干为产卵基质的卵孵化率最高,为87.41%,其次为完全腐烂的茎干,最后为正常茎干处理。

表1 二疣犀甲对不同腐烂状态椰子茎干产卵选择Table 1 Oviposition selection of Oryctes rhinoceros among coconut stem with different rotting state
2.2 对5种棕榈科植物的产卵选择
二疣犀甲对5种不同棕榈植物茎干的产卵选择结果如表2。从表中可知,二疣犀甲对不同棕榈植物茎干的产卵选择性不同。二疣犀甲在椰子和油棕茎干上的产卵量最高,单雌产卵量分别为24.28粒和21.22粒,两者之间差异不显著。而在槟榔、大王棕和蒲葵茎干上的产卵量显著少于椰子和油棕。
在不同棕榈科植物茎干基质中,二疣犀甲的卵孵化率有差异,其中以椰子茎干中的孵化率最高,为94.21%,与其它四种棕榈植物茎干处理相比均达到显著水平;其次为油棕茎干处理,孵化率为81.77%;大王棕和蒲葵茎干处理组孵化率最低,分别为46.50%和59.44%。
2.3 对不同死因椰子茎干的产卵选择
4种不同死亡原因造成的椰子茎干腐烂物对二疣犀甲的产卵诱集作用如表3。结果表明,二疣犀甲对不同原因造成的椰子死亡茎干产卵选择性具有明显差异,其中由台风等物理损伤引起的椰子死亡茎干对二疣犀甲的产卵引诱效果最强,产卵量为27.06粒;其次为染虫死亡茎干,产卵量为22.28粒;而感染泻血病和茎干腐烂病的椰子对二疣犀甲成虫的产卵引诱效果较差,产卵量分别为8.67和10.17粒。在孵化率统计中亦发现,二疣犀甲卵在台风物理损伤造成的死亡茎干基质中的孵化率最高,达到79.48%,其次为感虫死亡茎干基质,孵化率为57.98%,感病死亡的茎干基质处理组卵孵化率最低,且两种感病原因中孵化率差异不显著。

表3 二疣犀甲对不同死因的椰子茎干的产卵选择Table 3 Oviposition selection of Oryctes rhinoceros among coconut stem with different death cause
2.4 对5种椰子品种的产卵选择
二疣犀甲对不同品种椰子的茎干腐烂物产卵选择结果如表4。结果表明,二疣犀甲对不同品种椰子腐烂物上的产卵趋性存在差异。在供试5个品种的椰子茎干腐烂物中,以香水椰子对二疣犀甲的产卵引诱效果最强,单雌产卵量28.14粒,其次为海南高种椰子和红矮椰子,单雌产卵量分别为24.25和22.00粒,且这二者之间差异不显著。二疣犀甲在黄矮椰子和马哇椰子上产卵较少,产卵量分别为17.22和17.14粒,说明这两个品种的椰子对二疣犀甲的产卵引诱作用不强。
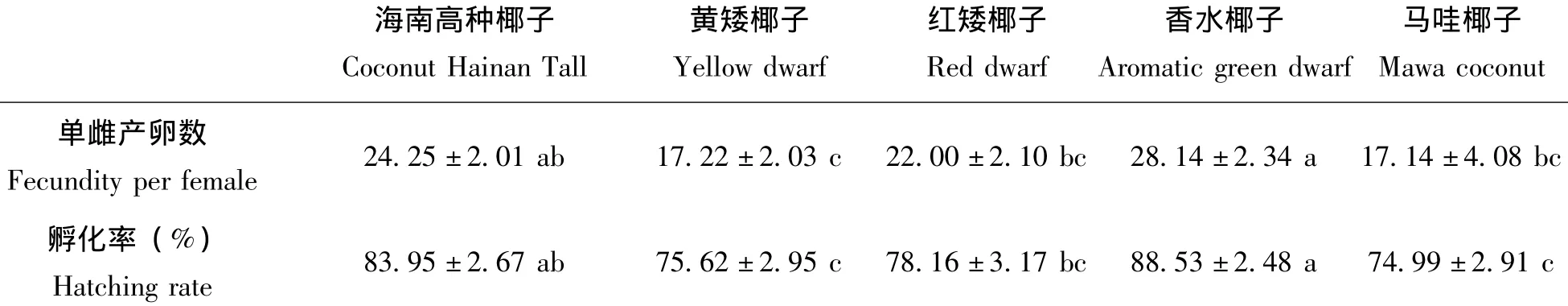
表4 二疣犀甲对不同椰子品种的产卵选择Table 4 Oviposition selection of Oryctes rhinoceros among different variety coconut stem
2.5 幼虫粪便对成虫产卵的影响
在半腐烂的椰子茎干中添加不同比例的幼虫粪便对二疣犀甲成虫产卵的影响如表5。由表可知:在产卵基质中添加一定量的幼虫粪便可以增加二疣犀甲成虫产卵量;当虫粪添加量达到产卵基质总量的25%时,二疣犀甲成虫产卵量和孵化率均最高,单雌产卵量达到34.67粒,孵化率为84.17%;之后随虫粪比例的继续增加,二疣犀甲产卵量呈下降趋势,其中在虫粪比例占产卵基质总量的55%时,单雌产卵量仅为15.78粒,孵化率为76.46%。
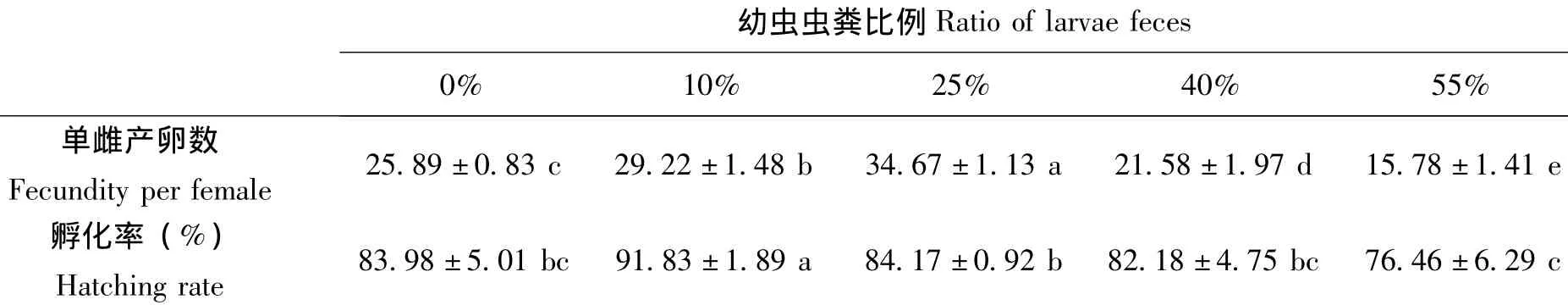
表5 幼虫粪便对二疣犀甲成虫产卵的影响Table 5 Effect of larvae dung on oviposition of Oryctes rhinoceros
3 讨论与结论
昆虫在对寄主利用的进化过程中,产卵选择性不仅受成虫产卵喜好与后代发育间联动性的影响,还存在其它影响因素 (Gratton and Welter,1998)。昆虫通过嗅觉识别寄主植物化学指纹图谱选择产卵寄主,当它与寄主植物直接接触后,还要依靠各种感受器对植物的化学和物理性状进行判断,检测其作为产卵寄主的适宜性 (张庆贺和姬兰柱,1994)。郝德君等研究表明松墨天牛对不同种类松树茎干的产卵选择具有差异 (郝德君等,2005),其中松墨天牛对黑松的选择性最强,对马尾松的选择性最弱,二者间差异显著。在黑松和马尾松的被松材线虫危害木与健康木之间,松墨天牛明显选择被害木产卵。具产卵痕枝段和虫粪处理过的枝段,对松墨天牛的产卵有显著抑制作用。
选择适宜的产卵场所是植食性昆虫生活史的重要环节之一,很多种类的初孵幼虫的活动力很差,必须依靠母体的适宜行为来发现可使其顺利生长发育的最佳食物来源。本研究选用南方栽种较为普遍的几种棕榈植物茎干、不同死亡程度及不同致死原因的茎干研究了二疣犀甲的产卵寄主选择特性,研究结果亦发现二疣犀甲对不同寄主的产卵选择具有差异,此结果不仅存在于不同棕榈种类之间,也存在于相同棕榈种类的不同品种内部 (不同椰子品种),这可能是因为不同寄主茎干内所含营养物质不同所致。田间调查发现,二疣犀甲仅选择死亡一段时间的棕榈茎干产卵,而对健康的棕榈茎干没有产卵取向,本研究中二疣犀甲在半腐烂和完全腐烂的茎干上产卵数量明显高于正常茎干,此结果与田间调查结果吻合,原因可能是茎干腐烂后内部组织发酵产生特殊气味等原因,也可能是腐烂后的椰子茎干的信息化合物产生了变化所致。在台风机械损伤和感染有害生物死亡的茎干中,二疣犀甲优先选择前者,究其原因可能是茎干被虫害和病害感染后,茎干内会残留虫害粪便、虫体分泌物、病菌等,这些物质会对二疣犀甲的产卵选择产生抑制作用。
由于植食性昆虫的生存与发育依赖于充足的植物食物资源,因此那些活动能力较差的种类必须进化一定的行为机理,以保证其不会在有限的食物资源上过分拥挤。郝德君等 (2005)研究发现虫粪处理过的枝段对松墨天牛具有产卵抑制作用,分析认为可能是虫粪含有产卵抑制物质。与郝德君等研究结果不同的是,本研究则发现一定量的虫粪比例会增加二疣犀甲的产卵选择几率,推测可能是因为二疣犀甲成虫具有通过产卵场所的粪便数量计算出此处幼虫数量的能力,如虫粪数量较少则说明幼虫数量较少,成虫就会继续选择此处产卵,当虫粪数量较多时则说明幼虫数量较多或可取食的食物较少,成虫即会重新选择产卵场所。
昆虫产卵行为的影响因素极其复杂,除了产卵基质状态和环境外,信息化学物质在调节昆虫产卵行为过程中起着十分重要的作用 (吴文伟等,2002)。收集可促使二疣犀甲成虫产卵的特殊茎干组织中的化学物质进行分离鉴定,并明确是寄主的挥发性次生代谢物质引诱二疣犀甲过来产卵,还是由于二疣犀甲接触到特定的化学物质后受到刺激致其产卵还有待进一步的研究。同时将分离出的活性成分深入研究,对于开发二疣犀甲新型引诱剂,以此来调控其产卵行为可为该虫的生态防控提供新的思路。
References)
Bedford G,1976.Mass rearing of the coconut palm rhinoceros beetle for release of virus.International Journal of Pest Management,22(1):5-10.
Cumber RA,1957.The rhinoceros beetle in Western Samoa.South Pacific Commission Technical Paper,107:76.
Dhileepan K,1988.Incidence and intensity of rhinoceros beetle infestation in oil palm plantations in India.Journal of Plant Crops,16:126-129.
Gratton C, WelterSC, 1998. Oviposition preferenceand larval performance of Liriomyza helianthi(Diptera:Agromyzidae)on normal and novel host plants. Environmental Entomology, 27(4):926-935.
Hao DJ,Zhang YJ,Dai HG,Wang Y,2005.Oviposition preference of Monochamus alternatus Hope(Coleoptera:Cerambycidae)to host plants.Acta Entomologica Sinica,48(3):460- 464. [郝德君,张永慧,戴华国,王焱,2005.松墨天牛对寄主树木的产卵选择.昆虫学报,48(3):460-464]
Mohd HT,Teoh CH,Kamarudzaman A,Mohd AA,1993.Zero burning- an environmentally friendly replanting technique.Proceedings of the 1993 PORIM International Palm Oil Congress.Agriculture Conference.Jalani BS,Ariffin D,Rajanaidu N,Mohd Tayeb D,Paramjothy K,Mohd BW,Henson IE,Chang KC,eds.PORIM,Bangi.185-195
Qin JD,1987.Relationship Between Insects and Plants:Review on Interaction between Insects and Plants and Its Evolution.Beijing:Science Press.38-61.[钦俊德,1987.昆虫与植物的关系—论昆虫与植物的相互作用及其演化.北京:科学出版社.38-61]
Sun JH,Roques A,Yan SC,2000.Behavioral manipulation methods and forest pest management.World Forestry Research,13(2):24-29.[孙江华,Roques A,严善春,2000.害虫行为调节与森林害虫管理.世界林业研究,13(2):24-29]
Wu WW,Chen JX,Song DL,2002.Progress in the study and application of ovipositioin deterring semiochemicals of insects.Southwest China Journal of Agricultural Sciences,15(3):105 -110.[吴文伟,陈建新,宋敦伦,2002.昆虫产卵忌避信息化学物质的研究及其应用.西南农业学报,15(3):105-110]
Zhang JP,Zhao BG,Wu ZH,Zhang YQ,2000.Application of behavior manipulation for pest management.Journal of Nanjing Forestry University,24(3):84-88.[张建平,赵博光,吴宗汉,张沂泉,2000.应用行为控制法治理害虫.南京林业大学学报,24(3):84-88]
Zhang QH,Ji LZ,1994.Chemical ecology of oviposition of herbivorous insects.Chinese Journal of Ecology,13(6):39-43. [张庆贺,姬兰柱,1994.植食性昆虫产卵的化学生态学.生态学杂志,13(6):39 -43]
猜你喜欢
南京林业大学学报(自然科学版)(2022年1期)2022-11-29
中国蚕业(2022年1期)2022-03-09
今日农业(2021年6期)2021-11-27
小天使·一年级语数英综合(2020年11期)2020-12-16
农业机械学报(2020年3期)2020-04-01
种子(2019年5期)2019-07-02
文理导航·科普童话(2017年4期)2018-02-10
散文诗(2017年17期)2018-01-31
风景园林(2017年3期)2017-06-22
天然产物研究与开发(2016年11期)2016-06-15
