
重组大肠杆菌产耐高温β-糖苷酶发酵条件优化
2013-03-19 02:12冯惠勇王立晖
河北工业科技 2013年1期
卢 楠,冯惠勇,王立晖
(1.河北化工医药职业技术学院制药工程系,河北石家庄 050026;2.河北科技大学生物科学与工程学院,河北石家庄 050018;3.天津现代职业技术学院生物化工系,天津 300222)
糖苷酶是一大类催化糖苷键水解的酶,在国际酶学命名方法中归位序EC 3.2.1,是目前用于酶法合成糖苷和低聚糖的一类重要的酶类。特别是某些种类的酶对乳糖有较高水解活性,在水解乳糖分子中β-1,4-半乳糖苷键的同时,具有转半乳糖基作用,以乳糖或其水解产物半乳糖和葡萄糖为糖基受体合成半乳糖苷键,生成低聚半乳糖(galacto-oligosaccharides,GOS)[1-10]。
本研究室构建保藏的基因工程菌株产生的耐高温β-糖苷酶ttβGLY具有较高的乳糖水解活性和转糖基活性,它的耐热性能有利于酶的分离纯化,可在高温体系中反应,具有反应速度快、转化率高的特点,在应用上有更大优越性。
笔者对本研究室构建保藏的产耐高温β-糖苷酶重组大肠杆菌的培养基和摇瓶发酵条件进行了探讨,为大幅度提高β-糖苷酶发酵产量提供依据。
1 材料与方法
1.1 材料
1.1.1 菌种
产酶菌株重组大肠杆菌,为本实验室构建菌种。
1.1.2 试剂
胰蛋白胨,酵母粉,生化试剂,北京奥博星生物技术有限公司提供;磷酸氢二钾,磷酸二氢钾,七水硫酸镁,乳糖,葡萄糖,蔗糖,甘油,尿素,氯化钠,皆为分析纯,天津市红岩化学试剂厂提供;氨苄青霉素,北京鼎国生物技术责任有限公司提供;葡萄糖试剂盒,北京北化康泰临床试剂有限公司提供;乳清,自制;玉米浆,石药集团维生药业有限公司提供;其他试剂均为市售分析纯或化学纯。
1.1.3 主要仪器
HH.BⅡ型电热恒温培养箱,天津市实验仪器厂提供;TGL-16C型台式离心机,上海安亭科学仪器厂提供;SW-CY-2FD型洁净工作台,上海博迅实业有限公司医疗设备厂提供;Multiskan MK3型酶标仪,上海雷勃分析仪器有限公司提供;HYG-ⅡB型生物反应摇瓶柜,上海欣蕊有限公司提供。
1.1.4 培养基
基础培养基为LB培养基:酵母粉(5g/L)、蛋白胨(10g/L)、氯化钠(10g/L),pH值调至7.0,1μL/mL加入100mg/mL的氨苄青霉素。
种子培养基:基础培养基为LB培养基,葡萄糖、碳酸钙按实验优化方案加入,pH值调至7.0,1μL/mL加入100mg/mL的氨苄青霉素。
发酵培养基:氯化钠(10g/L),碳源、氮源及其他成分按实验方案加入,pH值调至7.0,1μL/mL加入100mg/mL的氨苄青霉素。
1.2 实验方法
1.2.1 培养方法
种子培养:从斜面种子中取一环菌接种至装有50mL种子培养基的250mL三角瓶中培养,摇床转速为200r/min,温度为37℃,培养12h。
发酵培养:将培养好的种子培养液按照5%的接种量,接种至装有50mL发酵培养基的250mL三角瓶中进行发酵培养,摇床转速为200r/min,温度为37℃,培养24h。
每批实验都安排3组平行对照样,取实验结果的平均值进行分析和计算。
1.2.2 菌体量的测定
采用菌体干重法:发酵结束后,取适量菌液离心,弃去上清液,用蒸馏水洗涤后,将湿菌体放入烘干箱,于60℃下烘至恒重后测定菌体干重。
1.2.3 酶活的测定
酶活单位:以乳糖为底物,在温度70℃、pH值为7.0的条件下,将1min水解产生1mol葡萄糖定义为一个酶活单位(U)。
葡萄糖含量测定:采用葡萄糖氧化酶试剂盒法。
2 结果与讨论
2.1 摇瓶培养生长曲线和产酶曲线的测定
以LB为基础培养基,测定重组大肠杆菌的摇瓶生长曲线和产酶曲线,见图1。由图1可以看出:酶活和菌体均在培养时间为21~24h时达到较高,24h后随着时间延长,产酶量并没有进一步提高,因此确定24h为适宜的培养时间。由pH值变化曲线可以看出,在发酵过程中pH值从初始的7.0一直上升至8.6。这是由于LB培养基中含有大量氮源,而缺少碳源。碳氮比例的不均衡影响菌体的正常生长和酶活的表达,因此要以LB为基础培养基,添加不同的碳源及无机盐进行优化。

图1 菌体在LB基础培养基上的生长和产酶曲线Fig.1 Growth curve andβ-glucosidase production curve on LB medium
2.2 种子工艺的优化
2.2.1 葡萄糖加量的确定
初始培养基为LB培养基,鉴于葡萄糖为快速利用的碳源,有利于菌体的生长,此实验主要考察培养基中葡萄糖质量分数为0~1.0%对菌体生长的影响,并进行工艺优化。培养时间为12h,实验结果见表1。

表1 种子培养基中葡萄糖浓度对菌体生长的影响Tab.1 Effect of different glucose amount in seed medium on cell growth
由表1结果可知,葡萄糖质量分数为0.4%的菌体干重最高。但当葡萄糖质量分数为0.3%~1.0%时,发酵结束后pH值均低于7.0,而大肠杆菌的生长最适宜pH值为7.0,所以此时菌体生长受到抑制。
2.2.2 碳酸钙加量的确定
碳酸钙对发酵过程中的pH值具有缓冲作用。为了减少种子培养过程中pH值大幅度变化对菌体生长的不利影响,本实验考察了葡萄糖质量分数为0.4%时,添加碳酸钙对pH值和菌体生长情况的影响。结果见表2。
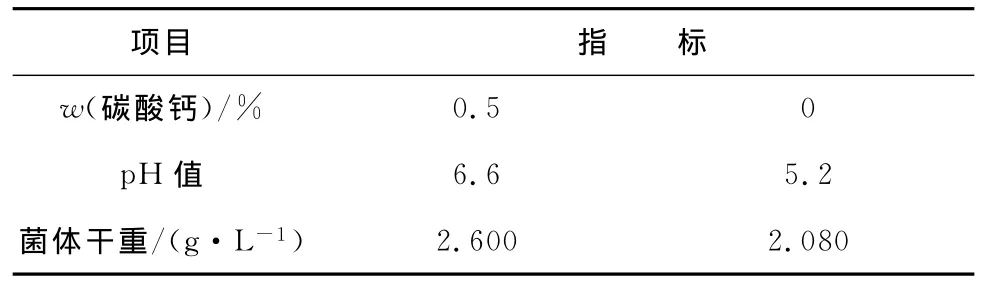
表2 种子培养基中碳酸钙加量对菌体生长的影响Tab.2 Effect of different CaCO3amount in seed medium on cell growth
由表2结果可知,加入0.5%(质量分数,下同)的碳酸钙后发酵液pH值变化幅度减小,更接近大肠杆菌生长的最适宜pH值。从菌体干重可看出,加入0.5%碳酸钙的菌体生长量更高。
2.2.3 种龄的确定
种龄在发酵过程中是一个重要影响因素。种龄短,种子启动慢,发酵时间延长;种龄长,菌种老化,代谢能力减弱。种龄一般选择在菌体生长的对数生长期的末期,本实验采用优化后的种子培养基,通过对种子生长曲线的测定,确定种子培养时间。结果见图2。
由图2可知,菌体干重在0~12h上升较快,之后变化较平缓,因此在优化后种子培养基条件下,选择12h为适宜的种子培养时间。
2.3 发酵培养基优化
2.3.1 碳源的选择
LB基础培养基主要成分为酵母浸粉、蛋白胨,含有丰富的氮源,但由于缺少适宜的碳源,因而在发酵过程中pH值持续上升,影响菌体的正常生长和酶活的表达。碳源物质不仅能促进重组菌的生长,而且对外源基因的表达有密切的影响。以LB培养基作为基础培养基,添加常用的5种碳源进行实验,其中乳清添加量以实际含乳糖的量折合计算。结果见表3。
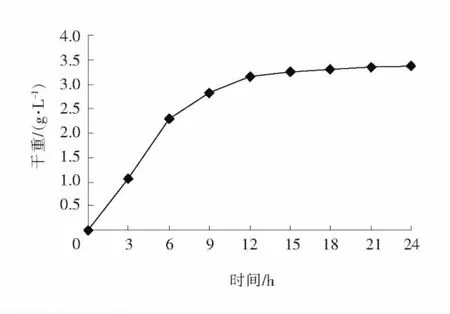
图2 菌体在优化后种子培养基上的生长曲线Fig.2 Growth curve on optimization of seed medium

表3 碳源对菌体生长及产酶的影响Tab.3 Effect of different carbon substrates on cell growth andβ-glucosidase production
由表3可知:以葡萄糖、蔗糖、甘油为碳源,干重、酶活比基础培养基均略有增长,但是pH值较低,pH值下降过快不利于酶活的表达和活性的保持;以乳糖、乳清为碳源,对菌体生长和酶活表达均有促进作用,干重和酶活增长较大,pH值变化较小,接近中性;乳糖、乳清添加量从1%增加到2%,菌体干重并没有明显变化,酶活反而降低。因此,综合考虑经济因素和产酶情况,选择乳糖和乳清以1%添加量作为碳源进一步优化。
2.3.2 氮源的选择
以6种氮源分别代替初始产酶培养基中的酵母粉和蛋白胨进行氮源单因素实验(分别以1%的乳糖和乳清为碳源),氮源添加量以氮源物质的量相等为依据。结果见表4。
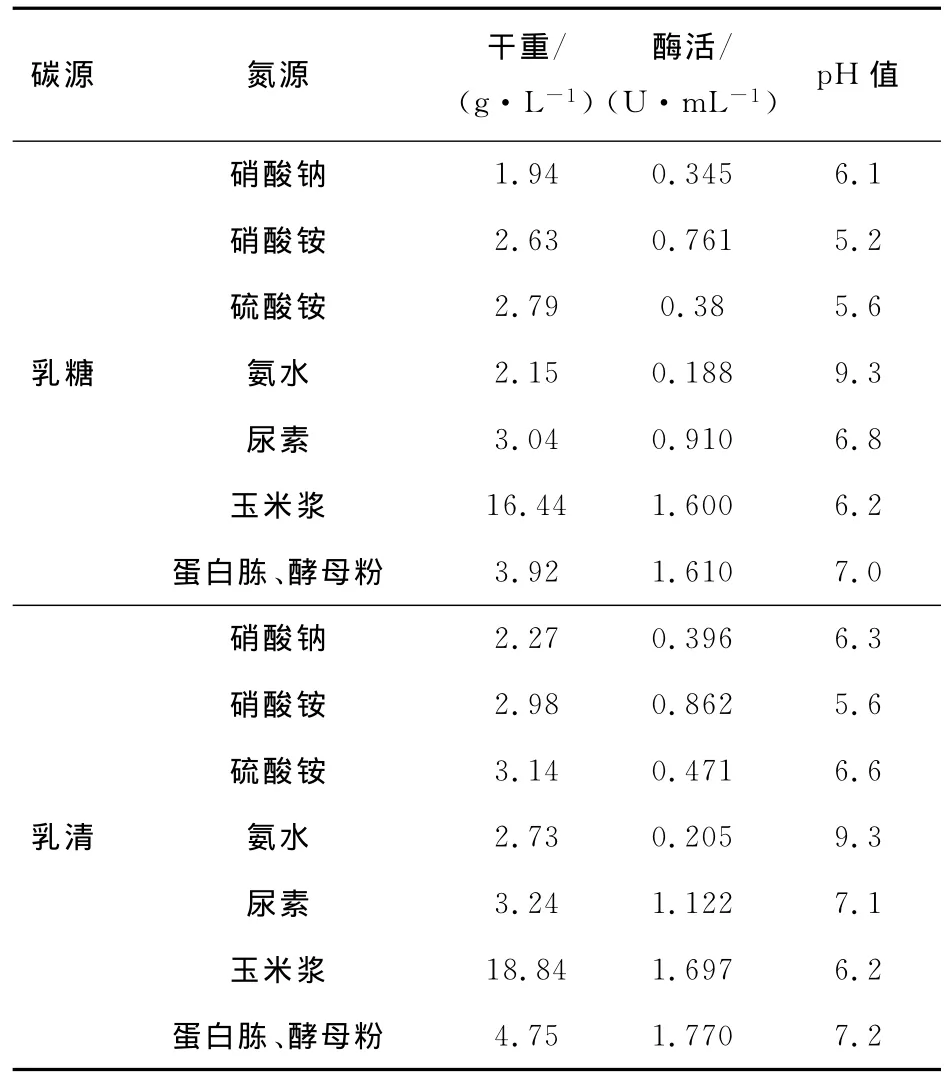
表4 氮源对菌体生长及产酶的影响Tab.4 Effect of different nitrogen substrates on cell growth andβ-glucosidase production
由表4可知:以乳糖和乳清为碳源的实验中,采用无机氮源时菌体量和酶活均低于以蛋白胨、酵母粉作氮源的结果;玉米浆为氮源时,酶活较高,但玉米浆中含有大量渣子、色素类物质,测得干重为16.44g/L,无法准确得出菌体的真正干重,且玉米浆作氮源发酵所制酶液由于含有色素类物质而导致固定化酶活较低。综合考虑,选用蛋白胨和酵母粉作为氮源。
分别以1%(质量分数,下同)的乳糖和乳清为碳源,以蛋白胨和酵母粉为氮源,改变蛋白胨和酵母粉的量,以确定合适的添加量。结果见表5。
由表5可知,以1%乳糖作为碳源,蛋白胨、酵母粉用量逐级增加,菌体干重随之增长,但增至蛋白胨1.2%、酵母粉0.6%后继续增加氮源酶活基本不变。以乳清作为碳源,蛋白胨、酵母粉用量逐级增加,菌体干重随之增长,蛋白胨1.4%、酵母粉0.7%后继续增加氮源酶活基本不变。考虑综合因素,分别选择碳源为1%乳糖,氮源为蛋白胨1.2%、酵母粉0.6%,记为L1培养基;碳源为1%乳清,氮源为蛋白胨1.4%、酵母粉0.7%,记为L2培养基。
2.3.3 无机盐配方优化
微生物要维持正常的生长代谢,除需要碳源和氮源外还需要Ca2+,Mg2+,K+等营养元素,这些元素将参与细胞的基础代谢,因此在配制培养基时必须以无机盐的形式加入各种必需离子。加入浓度过低,菌体生长缓慢;加入浓度过高,反而会抑制菌体生长,因此必须通过实验得出其合适的浓度范围。
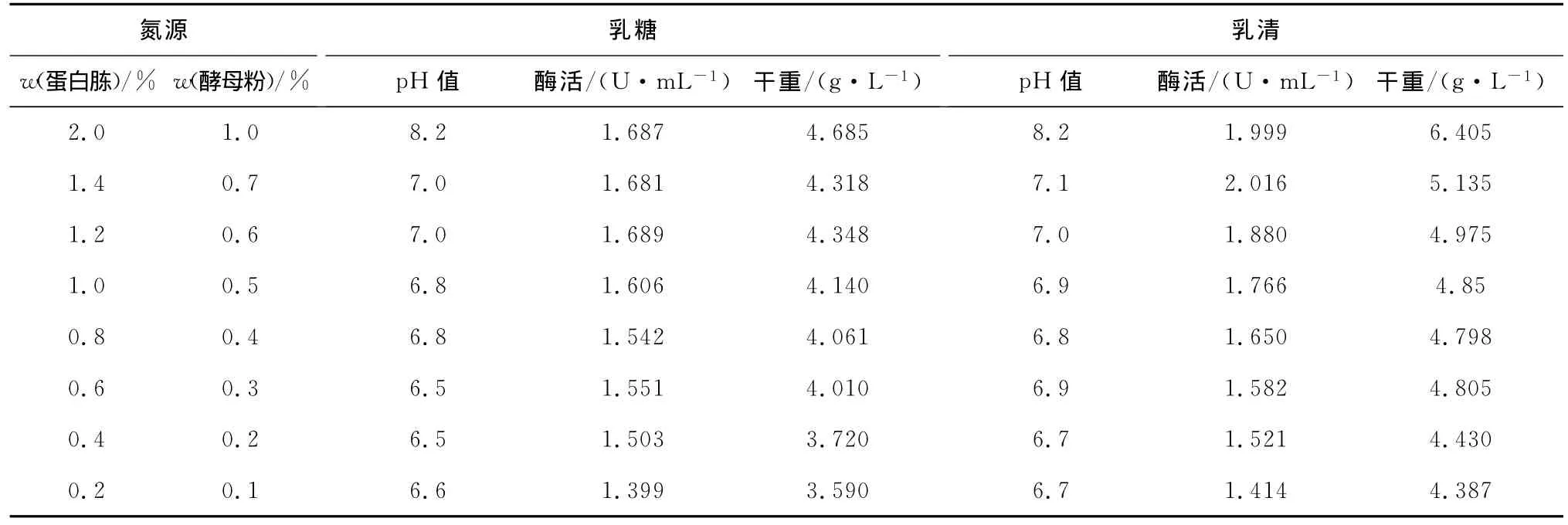
表5 氮源添加量对菌体生长及产酶的影响Tab.5 Effect of different nitrogen substrates amount on cell growth andβ-glucosidase production
常用无机盐有MgSO4·7H2O,MnSO4,KH2PO4,K2HPO4,KCl等。在L1和L2培养基中,分别添加KH2PO4和K2HPO4,考察磷水平对菌体生长和产酶的影响。结果见表6。

表6 磷添加量对菌体生长及产酶的影响Tab.6 Effect of different P amount on cell growth andβ-glucosidase production
由表6可知,L1和L2培养基添加磷盐量配比为KH2PO40.30%,K2HPO40.50%时有利于菌体的生长和产酶,把此配方培养基分别记为L3和L4培养基继续进行优化。
在L3和L4培养基中添加MgSO4·7H2O,考察Mg2+水平对菌体生长和产酶的影响。结果见表7。
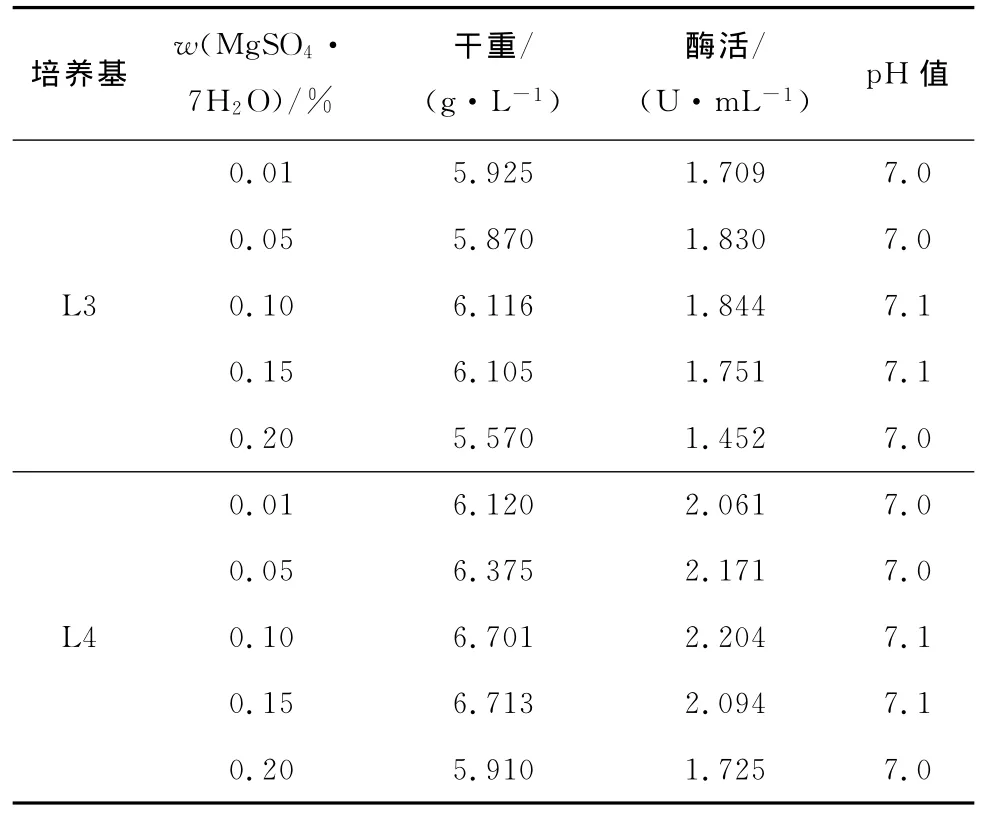
表7 Mg2+添加量对菌体生长及产酶的影响Tab.7 Effect of different Mg2+amount on cell growth andβ-glucosidase production
由表7可知,L3和L4培养基添加MgSO4· 7H2O为0.10%(质量分数)时有利于菌体的生长和产酶,把此配方培养基分别记为L5和L6培养基。
2.4 发酵条件的优化
发酵条件是影响重组菌生长与产酶的重要因素,以上述优化结果配制发酵培养基,对菌体产酶条件的影响因素进行考察,结果见表8。
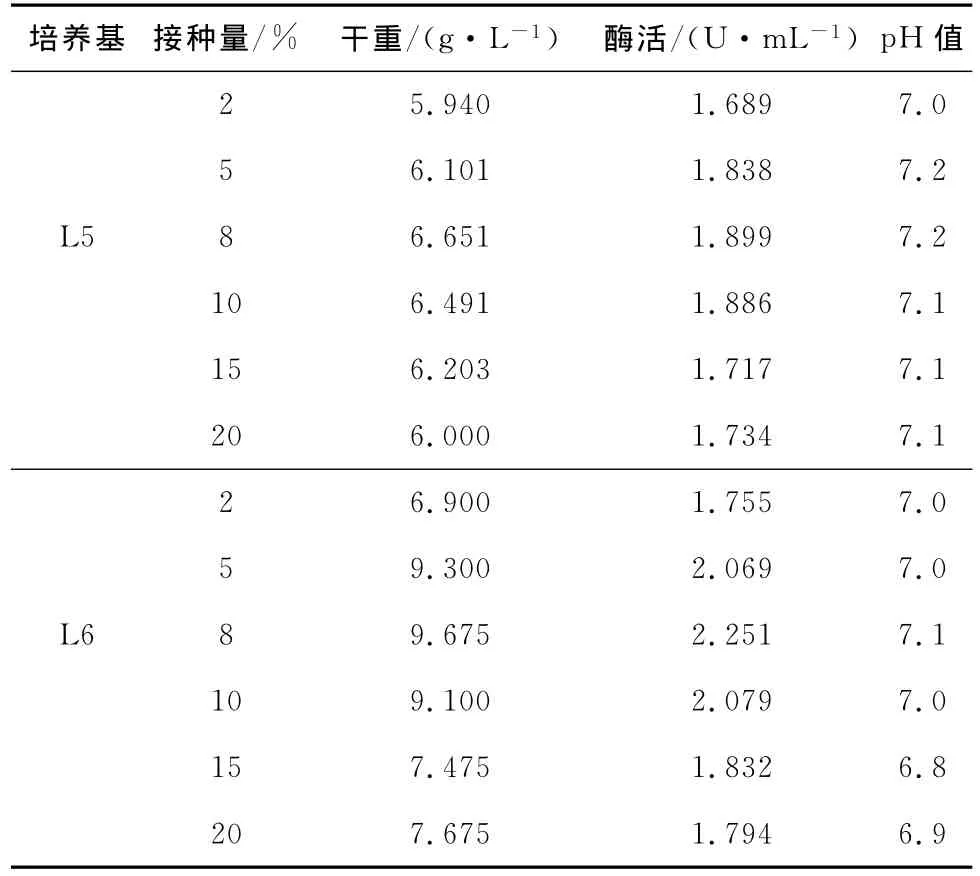
表8 接种量对菌体生长及产酶的影响Tab.8 Effect of different inoculum volume on cell growth andβ-glucosidase production
接种量的大小会直接影响发酵周期。接种量过小,发酵周期延长,增加成本;接种量过大,培养基内营养成分消耗快,不利于菌体生长和产酶。由表8可知,接种量为8%时,L5和L6培养基发酵的菌体干重和酶活均达到最高值,故选择8%为最适宜接种量。
在控制通气条件时必须考虑到既要满足菌体生长与胞内酶合成的不同需求,又要考虑节省能源,提高经济效益。在摇瓶培养条件下,可以通过改变摇床转速或者装液量来调节通气量。本研究固定的摇床转速为200r/min,通过改变三角瓶装液量来进行调解氧气的供应量,保持其他培养条件相同,在250 mL三角瓶中进行24h发酵,结果见表9。

表9 摇瓶装量对菌体生长及产酶的影响Tab.9 Effect of different amount of shaking flask on cell growth andβ-glucosidase production
由表9可以看出,当溶氧水平为250mL三角瓶中装液体培养基为40mL时,酶活最高。
3 结 论
对重组大肠杆菌发酵培养基进行了优化,确定出适合工程菌生长和耐高温β-糖苷酶表达的培养基配方:蛋白胨12g/L,酵母粉6g/L,乳糖10g/L,NaCl 10g/L,KH2PO43g/L,K2HPO45g/L,MgSO4·7H2O 1g/L。或采用乳清为碳源,培养基组成为蛋白胨14g/L,酵母粉7g/L,乳清13.3g/L,NaCl 10g/L,KH2PO43g/L,K2HPO45g/L,MgSO4·7H2O 1g/L。种子培养基组成为蛋白胨10g/L,酵母粉5g/L,葡萄糖4g/L,NaCl 10g/L,CaCO35g/L,种龄12h,接种量为8%,装量40mL/250mL三角瓶,于37℃发酵24h。优化后以乳糖为碳源,菌体干重、酶活最高可达6.830g/L和2.016U/mL,分别是优化前的3倍和2倍;以乳清为碳源,菌体干重、酶活最高可达10.010g/L和2.384U/mL,分别是优化前的4.4倍和2.4倍。
[1] FENG Huiyong,JULLIEN D,HOFFMANN L,et al.Converting aβ-glycosidase into aβ-thransglycosidase by directed evolution[J].JBC,2005,5:234-254.
[2] MICHEL D,LAURENT F,JEAN N L,et al.Cloning and expression of aβ-glycosidase gene from Thermus Thermophilus:Sequence and biochemical characterization of the encoded enzyme[J].Glycoconjugate Journal,1999(16):27-37.
[3] FOURAGE L,COLAS B.Synthesis ofβ-D-glucosyl-and-D-glucosyl-glucoses usingβ-glycosidase from Thermus thermophilus[J].Appl Microbiol Biotechnol,2001,56:406-410.
[4] 卢 楠,朱文华.谢氏丙酸杆菌发酵废液中生物抑菌物质的研究[J].河北工业科技,2012,29(3):138-140.
LU Nan,ZHU Wenhua.Study on biological antibacterial ma-terial in fermentation liquor of propionibacterium shermanii[J].Hebei Journal of Industrial Science and Technology,2012,29(3):138-140.
[5] 侯建革,王 芳,王丽丽,等.布氏乳杆菌产生甘露醇的发酵研究[J].河北科技大学学报,2010,31(2):132-136.
HOU Jiange,WANG Fang,WANG Lili,et al.Research in fermentation conditions of L-actobacillus buchneri CICC 6007 for product ion of mannitol[J].Journal of Hebei University of Science and Technology,2010,31(2):132-136.
[6] 陈 尧,孙国志,杨小荣,等.硅藻土助滤剂在头孢菌素C发酵液过滤中的应用研究[J].河北科技大学学报,2002,23(4):28-31.
CHEN Yao,SUN Guozhi,YANG Xiaorong,et al.A study on application of diatomide filter aid in the filtration of cephenlospr in C fermentation liquor[J].Journal of Hebei University of Science and Technology,2002,23(4):28-31.
[7] 付海云,王红权,何建勇.重组鲑鱼降钙素融合蛋白工程菌发酵工艺研究[J].河北科技大学学报,2006,27(1):54-56.
FU Haiyun,WANG Hongquan,HE Jianyong.Study on fermentation process of fusion protein of recombinant salmon calcitonin expressing E.coli[J].Journal of Hebei University of Science and Technology,2006,27(1):54-56.
[8] 赵 清,王 葭.三孢布拉霉的菌落形态与番茄红素发酵生产能力的关系[J].河北工业科技,2009,26(4):228-230.
ZHAO Qing,WANG Jia.Relationship between colonial morphology of blakeslea trispora and lycopene fermentation production capacity[J].Hebei Journal of Industrial Science and Technology,2009,26(4):228-230.
[9] 张 泉,郭晓宇,袁维峰,等.用于质粒DNA规模化生产的大肠杆菌发酵培养基的筛选[J].中国生物工程杂志,2006,27(1):102-105.
ZHANG Quan,GUO Xiaoyu,YUAN Weifeng,et al.Selection of liquid media for high density fermentation of recombinant E.coli for production of plasmids[J].China Biotechnology,2006,27(1):102-105.
[10] 杜木英,吴雪琴,侯大军,等.三种植物提取液的抑菌作用研究[J].四川食品与发酵,2003(3):5-8.
DU Muying,WU Xueqin,HOU Dajun,et al.Studies on the inhibitio lactivities of extracts of balsampear,garlie and capsicum[J].Sichuan Food and Fermentation,2003(3):5-8.
猜你喜欢
阅读(快乐英语中年级)(2022年5期)2022-06-03
江西水产科技(2019年3期)2019-09-10
天津农业科学(2019年4期)2019-05-31
山东工业技术(2016年15期)2016-12-01
中国酿造(2016年12期)2016-03-01
应用海洋学学报(2015年3期)2015-11-22
饲料博览(2014年7期)2014-04-05
食品工业科技(2014年15期)2014-03-11
食品工业科技(2014年9期)2014-03-11
中国食用菌(2013年2期)2013-11-21

- 河北工业科技的其它文章
- 冶金级硅水热酸洗提纯研究