
Gas7在成年大鼠脊髓和脊神经节的表达
2013-01-17 12:21侯良芹熊克仁丁艳霞
中国组织化学与细胞化学杂志 2013年3期
侯良芹 熊克仁 丁艳霞 丁 见
(皖南医学院人体解剖学教研室,安徽芜湖241000)
生长休止基因家族(growth arresting specific,Gas)是一类细胞培养过程中由于血清饥饿或者细胞接触性抑制时表达上调的基因,包括Gas1、Gas2、Gas3……Gas11等,这些基因编码的蛋白质缺少序列同源性,具有不同的功能[1-3]。Gas7是Gas基因家族中的一个亚型,主要表达在中枢神经系统的小脑、大脑皮层和海马,尤其高表达于小脑的浦肯野细胞[4]。资料显示,Gas7还表达于胚胎干细胞和发育过程中的神经系统,参与调控神经元的分化、发育和凋亡[5-7]。但Gas7在成年大鼠脊髓和脊神经节的表达和功能研究未见报道。本文采用RT-PCR方法、焦油紫染色以及免疫组织化学方法来检测Gas7基因核酸和蛋白在成年SD大鼠脊髓和脊神经节的表达,为进行Gas7功能研究提供参考资料。
材料和方法
1.实验动物及主要试剂
健康成年SD大鼠12只,清洁级,雌雄不拘,体重250±20g,购自南京青龙山动物中心,合格证号为SCXK(苏)2009-0001。6只用于 RT-PCR实验,6只用于免疫组化实验。
山羊抗Gas7抗体购自Santa Cruz公司;HRP标记的兔抗山羊二抗,DAB显色试剂盒均购自武汉博士德生物工程有限公司;Trizol溶液购自TaKa-Ra公司;Agarose-1000,Taq DNA Polymerase购自Invitrogen,USA。大鼠Gas7和β-actin引物由上海捷锐生物工程公司合成。
2.RT-PCR方法检测大鼠脊髓和脊神经节Gas7mRNA的表达
大鼠直接断头处死,咬骨钳打开椎管,取颈膨大部位的脊髓和与其相连的脊神经节0.1g,同时取小脑组织作为阳性对照。加入1ml Trizol溶液裂解。按Trizol试剂盒说明书提取总RNA。紫外分光光度计测总RNA浓度,取1μg的总RNA逆转录合成cDNA第一链待用,步骤按cDNA第一链合成试剂盒说明书进行。接着,以cDNA为模板进行PCR扩增,Gas7和内参β-actin的引物序列、PCR扩增片段的长度及PCR反应条件见表1。取PCR产物5μl,用浓度为1.2%的琼脂糖凝胶进行电泳,凝胶成像系统拍照。

表1 引物序列及PCR反应条件Table 1 Primers sequence and PCR condition
3.免疫组织化学方法检测Gas7蛋白在大鼠脊髓和脊神经节的表达
大鼠用剂量为50mg/kg的戊巴比妥钠深度麻醉后,经左心室先快速输入150ml 0.9%生理盐水,接着缓慢滴注含1%多聚甲醛和1.25%戊二醛的0.1mol/L磷酸缓冲液,结束后用咬骨钳打开椎管,取出脊髓和脊神经节,放入4%多聚甲醛磷酸缓冲液中固定12h,常规梯度酒精、二甲苯脱水、透明,石蜡包埋,连续冠状切片,片厚5μm,50℃烤片,时间长短依据组织切片与玻片的贴合情况而定,备用。
组织切片脱蜡入水后,依次经过微波抗原修复、3%H2O210min;5%BSA 37℃封闭30min后,滴加山羊抗Gas7抗体稀释液(1∶100),4℃冰箱过夜;滴加HRP标记的兔抗山羊二抗(1∶200),37℃孵育30min;DAB显色,显色时间长短依据镜下显色情况而定,5min左右;梯度酒精脱水(从75%开始),二甲苯透明,中性树胶封片,显微镜下观察并拍摄照片。本实验中,同时染色成年大鼠小脑组织切片作为阳性对照,用PBS代替一抗作为阴性对照。
4.焦油紫染色
为了更好地定位Gas7免疫阳性反应在脊髓的分布,我们将脊髓的切片同时进行了焦油紫染色,来显示脊髓神经元的分布和形态。步骤如下:石蜡切片常规脱蜡至水,在焦油紫染色液(由0.1%的焦油紫加少量冰醋酸制备而成)中染色,37℃温箱5-10min。梯度酒精分色(显微镜下观察分色程度,使尼氏小体呈深紫色而背景基本无色)、脱水透明,中性树胶封片。
结 果
1.Gas7mRNA在大鼠脊髓和脊神经节的表达
PCR结果显示,大鼠脊髓和脊神经节有较丰富的Gas7mRNA表达。在凝胶电泳图上(图1),脊髓和脊神经节Gas7的条带很清晰,稍弱于作为阳性对照的小脑组织的Gas7条带。

图1 RT-PCR结果琼脂糖凝胶电泳图,显示Gas7mRNA在成年大鼠脊髓和脊神经节以及小脑的表达。泳道1,小脑;泳道2,脊髓和脊神经节;泳道3,MarkerFig.1Electropherogram of Gas7mRNA expression in spinal cord and DRG,cerebellum of adult rat by RT-PCR on 1.2%agarose gel.Lane 1,cerebellum;Lane 2,spinal cord and DRG;Lane 3,Marker.
2.免疫组化方法显示Gas7蛋白在大鼠脊髓和脊神经节的表达和分布
Gas7蛋白免疫阳性为棕黄色颗粒。作为阳性对照的小脑组织切片上,浦肯野细胞呈强阳性着色(图2H),与以往的报道一致[4],而用PBS代替一抗作为阴性对照的切片上无阳性显色,说明本实验的结果是特异性的、准确可靠的。
在脊髓切片上,根据作为对照的焦油紫染色中神经元的分布,Gas7免疫阳性细胞的形态、大小和位置(图2A-C)如下:Gas7蛋白既表达于脊髓的灰质,也表达于白质。脊髓灰质前角的联络神经元和运动神经元均有表达,主要位于神经元的胞浆和胞膜,部分神经元的突起也呈阳性,细胞核不着色,尤其是Ⅸ版层大的α-运动神经元和小的γ-运动神经元,显色较深。颈膨大部位灰质前角Gas7阳性运动神经元数量较多(图2D),胸髓灰质前角数量较少(图2E)。脊髓灰质后角Ⅱ版层胶状质Gas7表达较强,阳性细胞多为体积很小的卵圆形神经元,分布较密集(图2F)。后角Ⅲ-Ⅵ版层Gas7阳性神经元形体较大,数量较少,散在分布。脊髓白质Gas7蛋白免疫阳性显色较弱且分布均匀(图2E)。脊神经节内大型感觉神经元呈Gas7免疫强阳性着色,中、小型感觉神经元为弱阳性着色,胞体为圆形或卵圆形(图2G)。
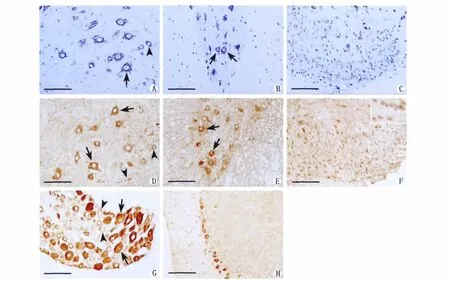
图2 脊髓组织切片(A颈膨大灰质的前角,B胸髓灰质的前角,C颈膨大灰质后角胶状质)的焦油紫染色以及Gas7蛋白在脊髓(D颈膨大灰质的前角,E胸髓灰质的前角,F颈膨大灰质后角胶状质)、脊神经节(G)和小脑(H)表达的免疫组织化学图片。图A、B、D和E,长箭头表示大的α-运动神经元,短箭头表示小的γ-运动神经元;图G,长箭头表示强阳性显色的大型感觉神经元,短箭头表示弱阳性显色的中、小型感觉神经元。图A-C是焦油紫染色;图D-H是免疫组织化学染色。标尺=100μm。Fig.2Nissl staining of adult rat spinal cord tissue section(A,B and C)and the expression of Gas7protein in adult rat spinal cord(D,E and F),DRG (G)and cerebellum (H)by immunohistochemistry.In A,B,D and E,arrow indicatedα-motor neurons and arrow head indicatedγ-motor neurons.In G,arrow indicated large diameter sensory neurons which were strongly stained and arrow head indicated medium and small diameter sensory neurons which were faintly stained.A-C:Nissl staining;D-H:Immunohistochemistry staining.Scale bars=100μm.
讨 论
目前的资料显示,Gas7基因主要表达于小脑、大脑皮层和海马等处[4],在神经系统其它部位的表达和功能的资料很少。本文用RT-PCR和免疫组化等方法研究了Gas7mRNA和蛋白在成年大鼠脊髓和脊神经节的表达和分布,发现脊髓和脊神经节有较高的Gas7mRNA的表达;脊髓灰质联络神经元和运动神经元以及脊神经节的感觉神经元都表达Gas7蛋白,主要位于神经元的胞浆、胞膜和突起,细胞核不表达。这些结果提示Gas7可能参与了脊神经节和脊髓感觉的传导和运动的控制。
脊神经节内的假单极神经元根据体积大小可分为大、中、小型3种,它们的周围突到达躯体和内脏的各种感受器,中枢突进入脊髓。其中大型神经元发出有髓鞘的粗纤维Aα和Aβ,进入脊髓后,部分纤维不在后角交换神经元,直接进入同侧脊髓后索,形成薄束和楔束传导本体感觉;部分纤维终止于后角,传导机械性感觉如触觉等。而中、小型神经元发出薄髓的Aδ和无髓的C纤维,主要终止于脊髓灰质后角的Ⅰ、Ⅲ、Ⅳ层,传导温度觉和痛觉等浅感觉[8]。大量传导痛觉和伤害性感觉的C纤维到达后角Ⅱ版层胶状质,胶状质由分布密集的小神经元组成,主要参与痛觉和伤害性感觉的分析[9-10]。本实验中,Gas7蛋白高表达于脊神经节的大型神经元,提示其对深感觉和机械性感觉的传导可能起着重要的作用。Gas7蛋白在脊神经节中、小型神经元的表达相对较弱,但在中、小型神经元发出的C纤维的主要靶区-后角胶状质,密集分布的小神经元高表达Gas7蛋白,说明其可能参与了痛觉的分析和调控。资料显示,Gas7蛋白位于胞浆和细胞膜附近,其羧基端序列能与肌动蛋白相互作用,促进肌动蛋白的聚合和肌动蛋白丝的交联,进而促进微丝微管的重组和形成,参与膜改变的动力学过程,例如突起的形成和细胞的迁移等[11-13]。本文观察到的Gas7蛋白表达于成年大鼠脊髓和和脊神经节的神经元,可能与神经元突起的维护和修复有关,进而调控神经元的功能,具体机制有待进一步的功能研究来阐明。
最近,Huang BT等人[14]为了研究 Gas7的生理功能,他们用转基因的方法制备了Gas7缺陷小鼠,发现随着年龄的增大,Gas7缺陷小鼠出现运动功能障碍。进一步的研究发现,Gas7缺陷小鼠脊髓运动神经元数量减少,骨骼肌快、慢肌纤维的构成发生改变,肌张力降低;同时,在神经肌接头处,神经纤维末端的出芽现象大大减少。本文的实验结果给出了合理的形态学上的解释,我们发现在脊髓灰质的前角,尤其是颈膨大和腰骶膨大处,Ⅸ版层内侧核和外侧核部位的躯体运动神经元高表达Gas7,包括大的α-运动神经元和小的γ-运动神经元。这些结果提示Gas7在运动神经元支配骨骼肌的生理过程中起着重要的调节作用,结合Gas7膜的动力学功能,可能参与了轴突末梢的生长和修复以及神经递质的释放等。
[1]Fabbretti E,Edomi P,Brancolini C,et al.Apoptotic phenotype induced by overexpression of wild-type gas3/PMP22:its relation to the demyelinating peripheral neuropathy CMT1A.Genes Dev,1995,9(15):1846-1856
[2]Li R,Chen J,Hammonds G,et al.Identification of Gas6as a growth factor for human Schwann cells.J Neurosci,1996,16(6):2012-2019
[3]Schneider C,King RM,Philipson L.Genes specifically expressed at growth arrest of mammalian cells.Cell,1988,54(6):787-793
[4]Ju YT,Chang AC,She BR,et al.gas7:A gene expressed preferentially in growth-arrested fibroblasts and terminally differentiated Purkinje neurons affects neurite formation.Proc Natl Acad Sci USA,1998,95(19):11423-11428
[5]Moorthy PP,Kumar AA,Devaraj H.Expression of the Gas7gene and Oct4in embryonic stem cells of mice.Stem Cells Dev,2005,14(6):664-670
[6]Hung FC,Cheng YC,Sun NK,et al.Identification and functional characterization of zebrafish Gas7gene in early development.J Neurosci Res,2013,91(1):51-61
[7]Hung FC,Chao CC.Knockdown of growth-arrest-specific gene 7b(gas7b)using short-hairpin RNA desensitizes neuroblastoma cells to cisplatin:Implications for preventing apoptosis of neurons.J Neurosci Res,2010,88(16):3578-3587
[8]Marmigère F,Ernfors P.Specification and connectivity of neuronal subtypes in the sensory lineage.Nat Rev Neurosci,2007,8(2):114-127
[9]Lu Y,Perl ER.A specific inhibitory pathway between substantia gelatinosa neurons receiving direct C-fiber input.J Neurosci,2003,23(25):8752-8758
[10]Hantman AW,van den Pol AN,Perl ER.Morphological and physiological features of a set of spinal substantia gelatinosa neurons defined by green fluorescent protein expression.J Neurosci,2004,24(4):836-842
[11]She BR,Liou GG,Lin-Chao S.Association of the growth-arrest-specific protein Gas7with F-actin induces reorganization of microfilaments and promotes membrane outgrowth.Exp Cell Res,2002,273(1):34-44
[12]Uchida T,Akiyama H,Sakamoto W,et al.Direct optical microscopic observation of the microtubule polymerization intermediate sheet structure in the presence of gas7.J Mol Biol,2009,391(5):849-857
[13]You JJ,Lin-Chao S.Gas7functions with N-WASP to regulate the neurite outgrowth of hippocampal neurons.J Biol Chem,2010,285(15):11652-11666
[14]Huang BT,Chang PY,Su CH,et al.Gas7-deficient mouse reveals roles in motor function and muscle fiber composition during aging.PLoS One,2012,7(5):e37702
猜你喜欢
冶金动力(2022年5期)2022-11-08
考试与评价·高二版(2020年2期)2020-09-10
中国临床医学影像杂志(2019年2期)2019-04-25
中国临床医学影像杂志(2019年2期)2019-04-25
鞍钢技术(2018年2期)2018-04-13
体育科技文献通报(2017年4期)2017-11-27
中国CT和MRI杂志(2014年7期)2014-06-27
中医研究(2013年9期)2013-03-11
中国医学科学院学报(2013年3期)2013-03-11
卒中与神经疾病(2012年1期)2012-10-18
