
电针对SAMP8小鼠海马线粒体呼吸链酶复合体活性的影响
2012-12-16 07:54董卫国陈采益林岚王丰谢永财陈跃
上海针灸杂志 2012年7期
董卫国,陈采益,林岚,王丰,谢永财,陈跃
(1.福建中医药大学中西医结合学院,福州 350108;2.福建中医药大学针灸学院,福州 350108)
阿尔茨海默病(Alzheimer’s disease,AD)是一种以进行性认知功能障碍和记忆损害为特征的中枢神经系统退行性疾病。AD好发于60岁以后的老年人,随着人口老龄化日益严重,AD发病率呈上升趋势[1]。但其病因及发病机制尚不十分清楚,目前没有特效治疗或逆转疾病进展的药物。针灸治疗AD具有多途径、多水平、多方式、多靶点的作用特点,而且疗效可靠,副反应少,越来越显示出其独特优势[2-4]。大量的证据表明,能量代谢障碍及线粒体结构及功能失调与 AD关系密切[5],而线粒体形态结构和功能的变化是导致 AD能量代谢障碍的重要因素和关键环节。本实验拟通过观察电针对快速老化小鼠亚系(senescence accelerated mouse/prone 8,SAMP8)海马神经元线粒体呼吸链酶复合体活性的影响,从线粒体功能角度进一步揭示针灸治疗AD的作用机制,为临床上针灸防治AD提供更充分的实验理论依据。
1 材料与方法
1.1 实验动物
8月龄SAMP8小鼠16只,8月龄抗快速老化小鼠亚系(senescence accelerated mouse/resistance1,SAMR1)8只,均为雄性,体重20~25 g,均由北京维通利华实验动物科技有限公司提供[实验动物质量合格证编号scxk(京)2002-0001]。
1.2 主要仪器及试剂
毫针(华佗牌无菌针灸针,规格 0.25 mm×15 mm),G6805电针治疗仪(上海医疗仪器厂),Morris水迷宫系统(中国医学科学院药物研究所),解剖显微镜(日本 Olympus公司),紫外分光光度计(Varian,美国),高效液相色谱仪系统(Waterz,美国)。提取及检测线粒体呼吸链酶复合体试剂,泛醌、氧化型 NADH、细胞色素C、抗霉素、EDTA、EGTA、DNA酶、Hepes、鱼藤酮、牛血清白蛋白(BSA)等均购自美国Sigma公司。
1.3 动物分组及治疗
动物在实验前适应性喂养1星期。采用计算机随机的方法将SAMP8鼠随机分为模型组、电针组,每组8只;SAMR1小鼠8只作为正常老化对照组。
电针组采用自制固定器固定小鼠,选择百会、大椎、肾俞(双侧)及太溪(双侧)进行电针治疗。具体方法为,直刺进针后,接G6805型电针治疗仪,选连续波,频率 2 Hz,电流强度以引起小鼠后肢肌肉轻微抖动而不嘶叫为宜,每次20 min,每日1次, 10 d为1个疗程,疗程间隔1 d进行下1个疗程,共3个疗程,计30 d。模型组和对照组不作治疗,只进行相同方式、相同时间和相同程度的抓取和小鼠固定器固定。
1.4 Morris水迷宫试验
采用 Morris水迷宫试验检测小鼠的空间学习记忆能力,测试内容包括定位航行试验和空间探索试验2部分。定位航行试验中,平台位于第3象限,没于水下1 cm,每天分上、下午两个时间段,每个时间段每鼠训练4次,将小鼠分别从4个入水点(各象限池壁中点)面向桶壁放入水中,记录小鼠爬上平台所需时间,即逃避潜伏期,如果90 s内未找到平台则将其引至平台并在平台停留 10 s,记录逃避潜伏期为 90 s,连续进行5 d。第6天撤离平台,进行空间探索试验,将小鼠从平台所在象限的对侧象限放入池中,自由游泳60 s后结束试验,记录小鼠在原平台象限停留的时间。
1.5 动物处理
水迷宫检测结束后,快速断头取脑,在冰浴上分离出完整海马,存放于-80℃冰箱,用于检测线粒体呼吸链酶复合体活性。
1.6 小鼠海马线粒体制备
整个制备过程均在0~4℃下进行。将取出的海马,称重,按 9:1(W/V)加入匀浆缓冲液(甘露醇210 mmol/L,蔗糖 70 mmol/L,三羟甲基胺基甲烷8 mmol/L,EGTA 1 mmol/L,pH 7.5)。匀浆 15次,于1300 g离心10 min,吸出上清液,将沉淀再次匀浆,离心后合并2次上清液,于1300 ×g离心10 min,收集上清液,于17000 ×g离心15 min,沉淀用匀浆缓冲液清洗2次,重悬于匀浆缓冲液中;以牛血清白蛋白为标准,采用Lowry法测定线粒体蛋白浓度,将其浓度调至200 μg/mL。
1.7 线粒体呼吸链酶活性测定
线粒体呼吸链酶复合体活性检测参照 Vyatlina的方法及结合我们以前的方法[6-7],用紫外分光光度法测定线粒体呼吸链复合体活性,将 10~20 μg的线粒体蛋白加入到终体积为2 mL的缓冲液中,以蒸馏水作空白管,校正光密度到0点,在30℃条件下,测定一定波长处 3 min内光吸收值的变化。酶活性单位为nmol/min/mg protein。
1.7.1 线粒体呼吸链酶复合体Ⅰ活性测定
将线粒体与 10 mM Tris- HCl(pH8.0)缓冲液、80 μmol/L泛醌、1 mg/mL BSA、0.25 mM铁氰化钾、和0.4 μmol/L抗霉素A混合后,加入200 μM NADH启动反应。连续测定340 nm处NADH吸收值的变化(NADH的消光系数ε=5.5 mmol/L-1cm-1)。
1.7.2 线粒体呼吸链酶复合体Ⅱ活性测定
将线粒体蛋白加入到复合物Ⅱ反应缓冲液内,混匀后加入20 μM琥珀酸盐启动反应。连续测定600 nm处 2,6-二氯靛酚钠盐水合物(DCPIP)吸收值的变化(DCPIP 的消光系数ε=19.1 mmol/L-1cm-1)。反应缓冲液含 50 mmol/L磷酸盐、pH 7.4,50 μmol/L 2,6-二氯靛酚钠盐水合物、2 mmol/L铁氰化钾、2 μg/mL鱼藤酮、2 μg/mL抗霉素A、25 μmol/L泛醌。
1.7.3 线粒体呼吸链酶复合体Ⅲ活性测定
将线粒体组分加入到线粒体呼吸链复合体Ⅲ缓冲液内,混匀后加入 80 μM还原型泛醌(DBH2)(以二硫苏糖醇还原)启动反应。连续测定550 nm处细胞色素C吸收值的变化(细胞色素C的消光系数ε=19.0 mmol/L-1cm-1)。反应缓冲液包括线粒体分离液、50 mmol/L Tris-HCl,pH 7.4、1 mmol/L EDTA、250 mmol/L蔗糖、2 mmol/L铁氰化钾、50 μmol/L氧化型细胞色素C。
1.7.4 线粒体呼吸链酶复合体Ⅳ活性测定
将线粒体蛋白加入到复合体Ⅳ反应缓冲液内,混匀后测定550 nm处细胞色素C吸光度变化3 min(细胞色素C的消光系数ε=19.0 mmol/L-1cm-1),即为细胞色素C氧化酶活性。缓冲液含线粒体分离液、10 mmol/L Tris-HCl pH 7.0,25 mmol/L蔗糖,120 mmol/L氯化钾,0.025%十二烷基β-D-麦芽糖苷,50 μmol/L还原型细胞色素 C。
1.8 统计学方法
数据以均值±标准差形式表示,采用SPSS13.0软件包进行统计学分析。定位航行试验采用重复测量数据方差分析,线粒体呼吸链酶复合体活性采用单因素方差分析(one way ANOVA),如差异有统计学意义,则用 LSD(least significant difference)法进行组间两两比较。P<0.05表示差异有统计学意义。
2 结果
2.1 各组小鼠逃避潜伏期比较
5 d的定位航行试验中,SAMR1对照组的逃避潜伏期明显短于SAMP8模型组(P<0.05或P<0.01),说明同一种系的正常老化小鼠学习记忆能力比快速老化小鼠强。与模型组比较,电针组平均逃避潜伏期缩短(P<0.05或P<0.01),并且第5天电针组平均逃避潜伏期接近对照组,提示电针能改善快速老化鼠的学习记忆能力。详见表1。
表1 各组小鼠平均逃避潜伏期比较 (±s,s)
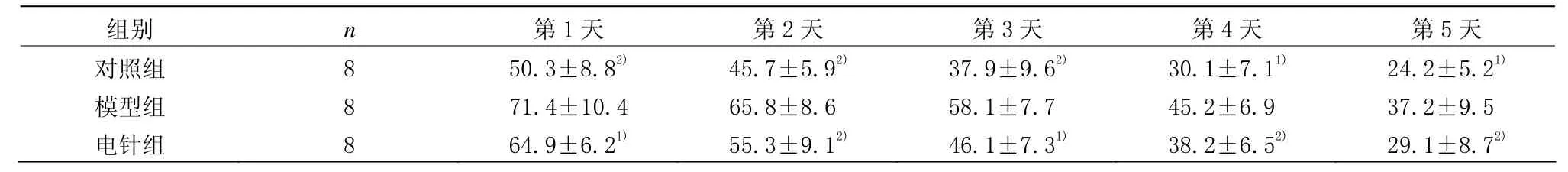
表1 各组小鼠平均逃避潜伏期比较 (±s,s)
注:与模型组比较1)P<0.05,2)P<0.01
组别 n 第1天 第2天 第3天 第4天 第5天对照组 8 50.3±8.82) 45.7±5.92) 37.9±9.62) 30.1±7.11) 24.2±5.21)模型组 8 71.4±10.4 65.8±8.6 58.1±7.7 45.2±6.9 37.2±9.5电针组 8 64.9±6.21) 55.3±9.12) 46.1±7.31) 38.2±6.52) 29.1±8.72)
2.2 各组小鼠在原平台象限停留时间的比较
表2 各组小鼠在原平台象限停留时间的比较 (±s,s)

表2 各组小鼠在原平台象限停留时间的比较 (±s,s)
注:与模型组比较1)P<0.05,2)P<0.01
组别 n 停留时间对照组 8 25.1±5.42)模型组 8 10.2±4.3电针组 8 17.3±6.21)
小鼠在原平台象限游泳时间越长,反映小鼠对原平台象限的记忆越好。模型组与 SAMR1对照组相比,时间明显缩短(P<0.01)。电针组时间较模型组延长(P<0.05),与对照组差异无统计学意义(P>0.05)。详见表2。
2.3 各组小鼠海马线粒体呼吸链酶复合体活性比较
对照组海马线粒体呼吸链酶复合体(复合体Ⅰ、Ⅱ、Ⅲ和Ⅳ)活性均较模型组明显增高(P<0.01)。电针组海马线粒体呼吸链酶复合体(复合体Ⅰ、Ⅱ、Ⅲ和Ⅳ)活性均较模型组明显升高(P<0.05)。电针组海马线粒体呼吸链酶复合体(复合体Ⅰ、Ⅱ、Ⅲ和Ⅳ)活性与对照组比较,差异无统计学意义(P>0.05)。详见表3。
表3 各组小鼠海马线粒体呼吸链酶复合体活性比较(±s,nmol/min/mg protein)
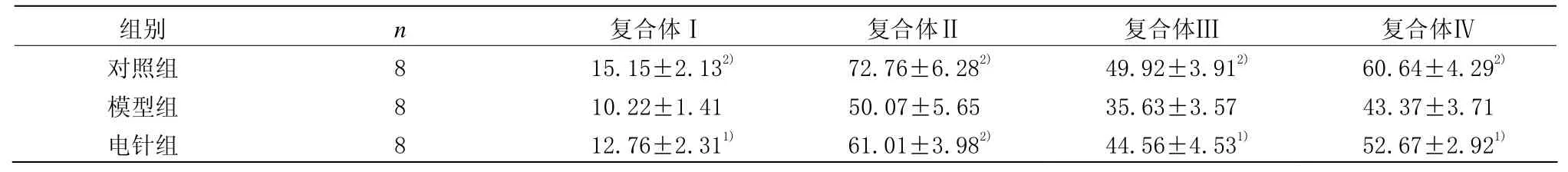
表3 各组小鼠海马线粒体呼吸链酶复合体活性比较(±s,nmol/min/mg protein)
注:与模型组比较1)P<0.05,2)P<0.01
组别 n 复合体Ⅰ 复合体Ⅱ 复合体Ⅲ 复合体Ⅳ对照组 8 15.15±2.132) 72.76±6.282) 49.92±3.912) 60.64±4.292)模型组 8 10.22±1.41 50.07±5.65 35.63±3.57 43.37±3.71电针组 8 12.76±2.311) 61.01±3.982) 44.56±4.531) 52.67±2.921)
3 讨论
线粒体是具有双层生物膜的细胞器,其主要功能是氧化代谢产物,并将产生的能量转化为 ATP,为机体提供能量。线粒体呼吸链酶复合体是位于线粒体内膜上与氧化磷酸化有关的蛋白质,主要包括复合体Ⅰ(NADH脱氢酶复合体),复合体Ⅱ(琥珀酸脱氢酶复合体),复合体Ⅲ(细胞色素还原酶复合体)及复合体Ⅳ(细胞色素氧化酶复合体)。线粒体呼吸链复合体活性能直接或间接反映线粒体的呼吸功能状况[8]。大量研究表明,AD的发生与线粒体密切相关,AD患者脑组织活检可见线粒体肿胀及数目减少[9]、DNA突变等形态结构的改变,还观察到线粒体膜电位降低,酶活性降低,ATP合成减少等功能异常。我们前期体外的研究结果也表明,β-淀粉样蛋白(β-Amyloid Protein,Aβ)诱导体外培养的海马神经元 AD模型可见线粒体呼吸链酶复合体活性降低,ATP含量减少[7]。这进一步说明了线粒体及线粒体呼吸链与AD的密切关系。本实验结果显示,与SAMP1对照组比较,SAMP8模型组海马线粒体呼吸链酶复合体活性降低。
SAMP8小鼠是由日本东京都大学竹田教授培育成功[10],SAMP8亚系的特征是中枢学习记忆功能随增龄进行性快速衰退[11]。研究表明,SAMP8脑内具有多种与AD类似的病理变化[12],因此,SAMP8小鼠被认为是一种相对较为完善的AD模型,已应用于AD的研究。所以,本实验中我们选择SAMP8作为AD动物模型进行研究。
Morris水迷宫是检测小鼠空间学习记忆的常用装置。定位航行试验检测小鼠的学习能力,而空间探索试验用来判断其近期记忆能力。通过实验发现,电针组潜伏期较模型组缩短,时间增加,表明电针可提高 AD模型小鼠的学习记忆能力,这与以前报道的结果一致。
已有学者研究报道,电针可减轻 SAMP8鼠海马神经元线粒体超微结构的损伤及提高线粒体细胞色素 C氧化酶活力[13-15],表明电针对线粒体的结构和功能都有保护作用。本实验从线粒体呼吸链酶复合体角度探讨电针治疗 AD的作用机制,其结果表明,电针能明显提高线粒体呼吸链酶复合体Ⅰ、Ⅱ、Ⅲ和Ⅳ活性,进一步证实了电针对线粒体功能的保护作用。由此,我们推测电针治疗 AD的机制可能是提高线粒体呼吸链酶复合体活性,从而改善线粒体功能及能量代谢。
[1] Walsh DM, Selkoe DJ. Deciphering the molecular basis of memory failure in Alzheimer's disease[J]. Neuron, 2004,44(1):181-193.
[2] 岳进,聂慧,李荣华.针灸治疗老年性痴呆的神经生物机制实验研究进展[J].上海针灸杂志,2009,28(3):183-185.
[3] Cheng H, Yu J, Jiang Z,et al. Acupuncture improves cognitive deficits and regulates the brain cell proliferation of SAMP8 mice[J].Neurosci Lett, 2008,432(2):111-116.
[4] Meng PY, Sun GJ, Mao JJ,et al. Effect of Electroacupuncture on Learning and Memory in Rats with Vascular Dementia[J]. J Acupunct Tuina Sci, 2012, 10(2):67-71.
[5] Moreira PI, Cardoso SM, Santos MS,et al. The key role of mitochondria in Alzheimer’s disease[J]. J Alzheimers Dis, 2006,9(2):101-110.
[6] Vyatkina G, Bhatia V, Gerstner A,et al. Impaired mitochondrial respiratory chain and bioenergetics during chagasic cardiomyopathy development[J]. Biochim Biophys Acta, 2004,1689(2):162-173.
[7] Dong W G, Huang F, Fan W G,et al. Differential effects of melatonin on amyloid-beta peptide 25-35-induced mitochondrial dysfunction in hippocampal neurons at different stages of culture[J]. J. Pineal Res, 2010,48(2):117-125.
[8] Rustin P, Chretien D, Bourgeron T,et al. Biochemical and molecular investigations in respiratory chain deficiencies[J]. Clin Chim Acta,1994,228(1):35-51.
[9] Hirai K, Aliev G, Nunomura A,et al. Mitochondrial abnormalities in Alzheimer’s disease[J]. J Neurosci, 2001,21(9):3017-3023.
[10] Takeda T, Hosokawa M, Takeshita S,et al. A new murine model of accelerated senescence[J]. Mech Ageing Dev, 1981,17(2):183-194.
[11] Takeda T. Senescence-accelerated mouse (SAM): a biogerontological resource in aging research[J]. Neurobiol Aging, 1999,20(2):105-110.
[12] Chen J, Zhou Y, Mueller-Steiner S,et al. SIRT1 protects against microglia-dependent amyloid-beta toxicity through inhibiting NF-kappaB signaling[J]. J Biol Chem, 2005,280(48):40364-40374.
[13] 彭静,曾芳,何宇恒,等.电针对SAMP8小鼠海马线粒体作用的研究[J].针刺研究,2007,32(6):364-367.
[14] 曾芳,何宇恒,彭静,等.电针对SAMP8小鼠海马神经元线粒体超微结构的影响[J].上海针灸杂志,2008,27(5):41-43.
[15] 何宇恒,曾芳,余曙光.电针对 SAMP8小鼠海马神经元线粒体超微结构及相关凋亡蛋白的影响[J].辽宁中医杂志,2008,35(10):160 0-1602.
猜你喜欢
中国科技纵横(2021年24期)2021-03-02
中成药(2018年6期)2018-07-11
原子与分子物理学报(2015年1期)2015-11-24
中国康复理论与实践(2015年7期)2015-05-09
中国中医药现代远程教育(2014年11期)2014-08-08
云南中医学院学报(2014年4期)2014-07-31
河南科技(2014年8期)2014-02-27
山西大同大学学报(自然科学版)(2013年5期)2013-09-13
食品科学(2013年15期)2013-03-11
化学分析计量(2013年3期)2013-03-11
