
NF-κB在抗β2GPⅠ/β2GPⅠ复合物诱导THP-1细胞表达组织因子中的作用探讨①
2012-11-28 02:03:34夏龙飞胡丽超陈东东解鸿翔
中国免疫学杂志 2012年11期
夏龙飞 周 红 胡丽超 陈东东 解鸿翔 王 婷 穆 原
(江苏大学基础医学与医学技术学院,镇江212013)
抗磷脂综合征(Antiphospholipid syndrome,APS)是一种以体内持续存在抗磷脂抗体(Antiphospholipid antibody,aPL)、临床以动静脉血栓形成、习惯性流产等为主要特征的自身免疫性疾病[1]。本组前期研究及相关文献显示,β2糖蛋白Ⅰ (beta-2-glycoproteinⅠ,β2GPⅠ)与其相应抗体(anti-β2GPⅠ)复合物(anti-β2GPⅠ/β2GPⅠ)能够刺激单核细胞、内皮细胞等表达组织因子(Tissue factor,TF),从而导致血液高凝状态[2]。进一步研究发现细胞表面受体AnnexinA2(ANX2)及Toll样受体4(Tolllike receptor 4,TLR4)作为β2GPⅠ的共受体,能够介导抗 β2GPⅠ/β2GPⅠ复合物对细胞的激活[3-5]。众所周知,TLR4被其配体激活后,将招募并激活细胞内一系列信号蛋白,如丝裂原活化蛋白激酶(Mitogen-activated protein kinases,MAPKs),最终导致相关基因的转录,从而引起相关效应[6,7]。核因子 κB(Nuclear factor-kappa B,NF-κB)是一种广泛存在的核转录因子,可参与对多种基因的转录来调控相关效应。近年来有研究显示NF-κB在TLR4相关的信号通路中发挥重要作用。本文主要探讨NF-κB在抗β2GPⅠ/β2GPⅠ复合物刺激THP-1细胞诱导TF表达过程中是否被激活及其作用。
1 材料与方法
1.1 主要材料及试剂 人单核细胞株THP-1(上海中国科学院细胞研究所),RPMI1640(Gibco公司),超级新生牛血清(Gibco公司),兔抗人NF-κB p65、磷酸化NF-κB p65单克隆抗体及兔抗人NF-κB抑制蛋白IκB-α多克隆抗体(Cell signaling公司),鼠抗人β-actin抗体(Protein tech Group公司),非特异性同型对照抗体R-IgG(武汉博士德生物工程有限公司),Trizol(Invitrogen公司),逆转录试剂盒(Toyobo公司),TF活性试剂盒(Assaypro公司),定量PCR试剂盒(杭州博日科技有限公司),ECL显色试剂(GE Healthcare 公司),LPS(Sigma 公司),NF-κB抑制剂PDTC、p38抑制剂SB203580(Sigma公司),ERK1/2抑制剂U0126(Promega公司),JNK抑制剂SP600125(R&D公司),多克隆抗β2GPⅠ抗体和β2GPⅠ(本实验室制备),引物(设计采用primer 5.0,由上海生物工程有限公司合成),其他试剂均为国产分析纯。
1.2 方法
1.2.1 THP-1细胞培养 采用 RPMI1640培养液(含10%新生牛血清,100 U/ml青霉素和100 U/ml链霉素)培养THP-1细胞,置于37℃、5%CO2饱和湿度的孵育箱培养,待细胞长满培养瓶底部70% ~80%时,细胞传代,取传代3~10次的细胞用于实验。
1.2.2 细胞总RNA的提取和逆转录 将状态良好的THP-1细胞种入6孔板内(2×106/孔),37℃、5%CO2培养24小时后,弃去培养液,换1 ml无血清RPMI1640继续培养12小时。根据需要将细胞与NF-κB抑制剂PDTC(20 μmol/L)预先孵育90分钟,然后根据实验设计加入不同刺激物处理细胞2小时,收集细胞,加入1 ml Trizol裂解,混匀后于室温放置20分钟充分裂解,按Trizol说明书提取细胞总RNA。逆转录体系按照试剂盒说明操作,反应条件:37℃ 30分钟,98℃ 5分钟,4℃ 5分钟。反应所得cDNA用于检测mRNA表达。
1.2.3 细胞裂解物的制备 收集传代3~10次、状态良好的THP-1细胞,种于6孔板内,密度为2×106/孔,37℃、5%CO2培养箱培养24小时后,弃去上清,加入无血清RPMI1640培养液饥饿12小时后,弃上清,再加1 ml无血清RPMI1640培养基,然后用不同刺激物处理细胞。抑制试验时将不同抑制剂(SB203580,10 μmol/L;SP600125,5 μmol/L;U0126,90 nmol/L)与细胞预处理30分钟再加刺激物。收集细胞,加入适量细胞裂解液,于冰上裂解1小时,期间每隔15分钟剧烈震荡一次,4℃ 11 200 r/min离心10分钟后取上清,测定蛋白浓度,-70℃保存备用。
1.2.4 实时定量 PCR(Real time quantitative PCR,RT-qPCR)检测 定量PCR分析仪检测细胞TF mRNA水平,总反应体系为12 μl:其中2×SYBR Mix 6 μl,上下游引物各0.48 μl,最后用灭菌 ddH2O补足至12 μl。TF引物序列为:上游5'-TCAGGTGATCCACCCACCTT-3',下 游 5'-GCACCCAATTTCCTTCCATTT-3',扩增产物为211 bp;β-actin引物序列为:上游 5'-CACGAAACTACCTTCAACTCC-3',下游5'-CATACTCCTGCTTGCTGATC-3',扩增产物为262 bp。RT-qPCR循环参数为:95℃预变性5分钟;一个循环:95℃ 15 秒,62℃(β-actin、TF)20 秒,72℃20秒;38个循环扩增。目的基因的相对表达水平=目的基因拷贝数/β-actin拷贝数。
1.2.5 Western blot分析 取上述收集的细胞裂解物标本(总蛋白60 μg)与样本缓冲液作用,94℃变性5分钟后,12%SDS-PAGE电泳分离蛋白,再以350 mA恒流转印至PVDF膜;膜置于5%脱脂牛奶的TBS/T室温封闭1小时;再分别用相应的兔抗人NF-κB p65(1∶1 000)、p-NF-κB p65(1∶1 000)、IκB-α抗体或鼠抗人β-actin抗体(1∶2 500)4℃孵育过夜;次日用TBS/T洗涤3次,15分钟/次,采用HRP标记的羊抗兔 IgG(1∶2 000)或羊抗鼠 IgG(1∶3 000),37℃孵育 1小时,TBS/T洗涤 3次,15分钟/次,ECL显色系统定影显色,观察杂交条带,同时运用Champchemi分子成像系统扫描条带并读取灰度值。
1.2.6 TF活性检测 利用TF/Ⅶa复合物使因子Ⅹ转变为活化因子Ⅹa,根据Ⅹa生成量来判定细胞TF的活性。测定标本为细胞裂解物,严格按照试剂盒说明书操作。在96孔反应板中加入50 μl的样品稀释液和10 μl的因子Ⅶ后,分别加入倍比稀释的标准品和测定样本10 μl,以样品稀释液为空白,37℃反应30分钟后,加入因子Ⅹ。37℃反应30分钟后再加入Ⅹa的发色底物,于405 nm、37℃ 条件下测定吸光度值。吸光度值与样品中TF活性呈正相关,TF活性结果以pmol/L(pmol/L)表示。
1.3 统计学分析 采用软件SPSS10.0进行统计学分析,所有数据以±s表示。对单因素处理组之间数据的比较采用One-way ANOVA方法,对双因素处理组之间数据的比较采用Two-way ANOVA方法,P<0.05为差异具有统计学意义。
2 结果
2.1 抗β2GPⅠ/β2GPⅠ复合物诱导THP-1细胞表达TF mRNA及活性 前期研究发现抗β2GPⅠ/β2GPⅠ复合物能够诱导血液单核细胞及单核细胞株THP-1细胞表达 TF。本实验以 R-IgG/β2GPⅠ(同型抗体对照,100 μg/ml)、抗 β2GPⅠ/BSA(同型抗原对照,100 μg/ml)、LPS(阳性对照,500 ng/ml)及抗 β2GPⅠ/β2GPⅠ复合物(100 μg/ml)分别刺激THP-1细胞2、6小时后,分别收集细胞提取总RNA和细胞裂解物,进行了RT-qPCR及TF活性试剂盒分析。如图1显示,抗β2GPⅠ/β2GPⅠ 复合物使TF mRNA表达水平显著增加及TF活性明显增强,与LPS具有相同的刺激效应,与空白对照组(media)比较,差异显著(P<0.05),而同型抗体对照R-IgG/β2GPⅠ和抗原对照抗β2GPⅠ/BSA则无此作用。
2.2 抗 β2GPⅠ/β2GPⅠ复合物促进 THP-1细胞NF-κB的活化 为了观察 NF-κB是否在 anti-β2GPⅠ/β2GPⅠ刺激THP-1细胞过程中被激活,我们收集刺激不同时间点的细胞总蛋白,利用相应抗体进行Western blot分析。由图2可见,THP-1细胞经anti-β2GPⅠ/β2GPⅠ复合物刺激后,NF-κB 总蛋白(t-NF-κB p65)水平变化不明显,但是NF-κB 的磷酸化(p-NF-κB p65)在逐渐增强,并在90分钟时达到高峰而后降低;另外,与NF-κB的磷酸化相反,IκB-α蛋白水平随复合物作用逐渐下降,于90分钟达到最低水平,之后逐渐恢复。说明抗β2GPⅠ/β2GPⅠ复合物作用于细胞,促进了 IκB-α 与 NF-κB(p65/Rel A)解离,后者被激活从而发挥转录活性。
2.3 不同刺激物对THP-1细胞NF-κB p65磷酸化及IκB-α 蛋白的影响 上述结果表明,anti-β2GPⅠ/β2GPⅠ能够促进THP-1细胞内NF-κB磷酸化并具有时效性,本实验进一步验证这一效应的特异性。我们采用了不同组合刺激物处理THP-1细胞90分钟,收集细胞提取蛋白标本,进行Western blot分析,比较各组的刺激效应。结果显示,同源对照抗体与β2GP Ⅰ组合(R-IgG/β2GP Ⅰ,100 μg/ml)以及无关抗原对照抗牛血清白蛋白与β2GPⅠ(anti-β2GPⅠ/BSA,100 μg/ml)均不能引起 NF-κB p65的磷酸化以及IκB-α 降解;而抗 β2GPⅠ/β2GPⅠ (100 μg/ml)能够明显增加磷酸化NF-κB p65的表达(图3),与空白对照(media)比较,条带明显增强,同时降低IκB-α蛋白含量,条带弱于对照;LPS(500 ng/ml)作为阳性对照,具有增强 NF-κB p65磷酸化并降低 IκB-α含量的效应。表明anti-β2GPⅠ/β2GPⅠ复合物刺激THP-1细胞NF-κB p65磷酸化的刺激效应具有特异性,结果与LPS相似。

图1 Anti-β2GPⅠ/β2GPⅠ诱导的 THP-1细胞 TF mRNA(A)表达及TF活性(B)Fig.1 Anti-β2GPⅠ/β2GPⅠ complex induces TF mRNA expression(A)and TF activity(B)in THP-1 cells
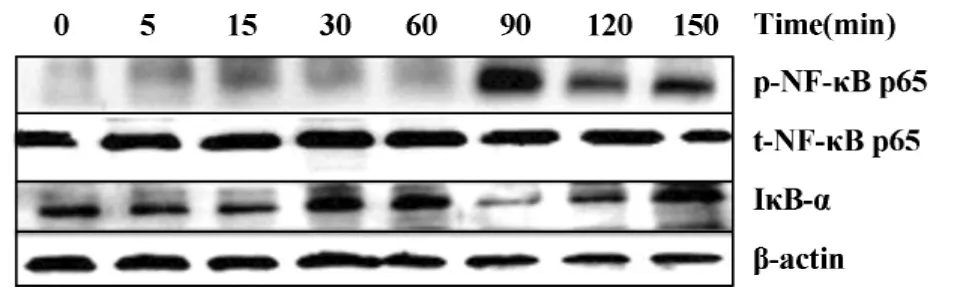
图2 Anti-β2GPⅠ/β2GPⅠ刺激 THP-1 细胞 NF-κB p65的磷酸化及IκB-α变化的时效性Fig.2 Time course of anti-β2GPⅠ/β2GPⅠ-induced NF-κB p65 phosphorylation and IκB-α change in THP-1 cells
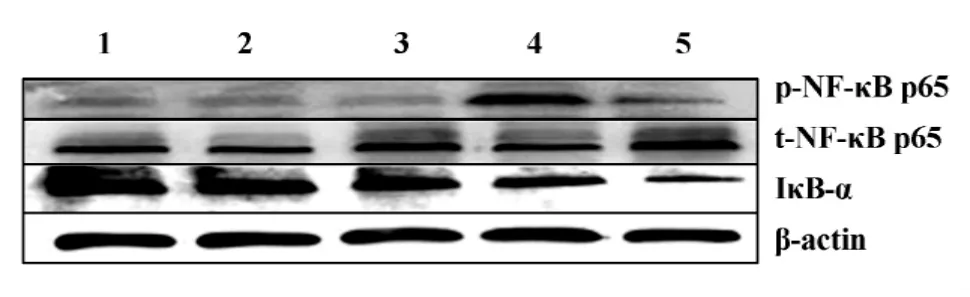
图3 Anti-β2GPⅠ/β2GPⅠ刺激 THP-1 细胞 NF-κB p65磷酸化及IκB-α变化的特异性Fig.3 Specific effects of anti-β2GPⅠ/β2GPⅠon NF-κB p65 phosphorylation and IκB-α change in THP-1 cells
2.4 NF-κB 抑制剂 PDTC 对 anti-β2GPⅠ/β2GPⅠ诱导THP-1细胞TF表达的干预效应 本实验进一步明确了NF-κB抑制剂PDTC是否影响anti-β2GPⅠ/β2GPⅠ诱导THP-1细胞TF表达的作用。结果显示:PDTC(20 μmol/L)能够明显降低 anti-β2GPⅠ/β2GPⅠ刺激细胞表达TF mRNA(图4A)和TF活性(图4B),与PDTC未处理组相比,差异具有统计学意义(P<0.05),而PDTC对无刺激的THP-1细胞没有明显影响(P>0.05)。本实验进一步表明NF-κB 在anti-β2GPⅠ/β2GPⅠ复合物诱导 THP-1 细胞表达TF的过程中具有重要的作用。另外,NF-κB抑制剂PDTC也能够抑制LPS对细胞TF表达的刺激效应。
2.5 上游信号分子MAPKs的抑制剂对抗β2GPⅠ/β2GPⅠ复合物诱导THP-1细胞NF-κB磷酸化的影响 为明确在抗β2GPⅠ/β2GPⅠ复合物诱导THP-1细胞表达TF过程中NF-κB的磷酸化与信号通路中上游关键分子 MAPKs之间的关系,我们采用了MAPKs抑制剂进行试验。结果显示:p38抑制剂SB203580、ERK1/2抑制剂 U0126及 JNK抑制剂SP600125分别单独作用细胞,可使抗β2GPⅠ/β2GPⅠ刺激细胞NF-κB磷酸化水平有所降低,与无抑制剂处理组比较,p-NF-κB p65条带减弱(图5),对LPS刺激细胞NF-κB磷酸化似乎无明显影响;若同时使用二种抑制剂(SB203580/U0126,U0126/SP600125,SB203580/SP600125),均可使抗 β2GPⅠ/β2GPⅠ及LPS引起的NF-κB磷酸化水平明显降低,条带显著减弱(图5)。三种抑制剂对未刺激的THP-1细胞基础NF-κB磷酸化水平无抑制作用(见media组)。

图4 NF-κB特异性抑制剂PDTC干预anti-β2GPⅠ/β2GPⅠ诱导的THP-1细胞TF mRNA表达(A)及TF活性(B)Fig.4 The effects of NF-κB inhibitor PDTC on anti-β2GPⅠ/β2GPⅠ-induced TF mRNA expression(A)and TF activity(B)in THP-1 cells

图5 MAPKs抑制剂对anti-β2GPⅠ/β2GPⅠ处理THP-1细胞对NF-κB活化的影响Fig.5 The effects of MAPKs inhibitors on anti-β2GPⅠ/β2GPⅠ-induced NF-κB activation in THP-1 cells
3 讨论
核因子κB(NF-κB)是从B淋巴细胞核抽提物中检测到的一种能与免疫球蛋白κ轻链基因增强子κB序列特异结合的核蛋白因子,是一类普遍存在的转录因子。哺乳动物NF-κB家族包括NF-κB1(p50)、NF-κB2(p52)、RelA(p65)、RelB 和 c-Rel五个成员,除了Rel-B只能与P50或者P52有效的结合外,存在所有的同源或异源二聚体组合的可能性,并且都具有NF-κB的活性,最常见的二聚体形式是p65/p50异源二聚体。静息状态下NF-κB二聚体与IκB结合,以无活性形式存在于胞浆。而当TNF-α、TLR激动剂等刺激信号作用细胞时,促使IκB-α磷酸化,然后被泛素结合酶识别发生快速泛素化继而被蛋白酶体蛋白水解。随着IκB-α的降解,NF-κB二聚体被释放,进而迅速转入核内,与相应的κB位点结合,从而对相关基因表达进行调控[8]。NF-κB体系主要涉及机体防御反应、组织损伤和应激、细胞分化和凋亡以及肿瘤生长抑制过程的信息传递[9]。
本研究小组前期对抗磷脂综合征(APS)血栓形成机制的探讨,证明了抗β2GPⅠ/β2GPⅠ复合物刺激血液单核细胞表达TF是其主要机制之一。并发现抗β2GPⅠ/β2GPⅠ复合物能够产生与LPS相同的刺激效应,显著增加单核细胞株 THP-1表达TLR4、MD-2及MyD88 mRNA及蛋白水平的表达,并且部分依赖ANX2,随后招募一系列蛋白(包括IRAK1/4、TRAF6及MAPKs)发生级联反应,最终促进细胞表达TF,从而参与APS血栓形成[10]。本文进一步发现NF-κB在抗β2GPⅠ/β2GPⅠ复合物诱导THP-1细胞表达TF过程中被诱导活化,提示NF-κB在其中扮演着重要作用。近年来有文献报道NF-κB参与了抗磷脂抗体(aPL)诱导内皮细胞及单核细胞活化[11,12],但是关于 NF-κB 参与抗 β2GPⅠ/β2GPⅠ复合物诱导THP-1细胞表达TF鲜有报道。本研究采用不同对照刺激物处理细胞后,发现抗β2GPⅠ/β2GPⅠ对NF-κB的活化具有特异性。
为进一步证明 NF-κB在抗 β2GPⅠ/β2GPⅠ诱导THP-1细胞表达TF过程中的作用,本研究采用NF-κB特异性抑制剂PDTC以及上游关键信号分子MAPKs特异性抑制剂(p38、ERK及JNK的特异抑制剂SB203580、U0126、SP600125)进行验证。结果显示PDTC能够抑制抗β2GPⅠ/β2GPⅠ诱导THP-1细胞TF的表达,并且MAPKs特异性抑制剂能够抑制NF-κB的活化。有文献报道MAPKs的活化对于抗β2GPⅠ/β2GPⅠ刺激细胞表达相关因子是必需的,结合本文结果,进一步表明在抗β2GPⅠ/β2GPⅠ诱导 THP-1细胞表达TF的信号转导过程中,MAPKs/NF-κB通路至关重要,提示可作为分子靶点用于防治抗磷脂综合征的血栓形成[7,8]。
1 Zhou H,Wolberg A S,Roubey R A.Characterization of monocyte tissue factor activity induced by IgG antiphospholipid antibodies and inhibition by dilazep[J].Blood,2004;104(8):2353-2358.
2 Sorice M,Longo A,Capozzi A et al.Anti-beta2-glycoprotein I antibodies induce monocyte release of tumor necrosis factor alpha and tissue factor by signal transduction pathways involving lipid rafts[J].Arthritis Rheum,2007;56(8):2687-2697.
3 Zhou H,Ling S C,Yu L et al.Involvement of annexin A2 in anti-β2GPⅠ/β2GPI-induced tissue factor expression on monocytes[J].Cell Res,2007;17:737-739.
4 严一红,周 红,周保成 et al.TLR4在anti-β2GPI/β2GPI复合物诱导THP-1细胞表达TF中的作用探讨[J].中国免疫学杂志,2010;26(5):396-401.
5 Zhou H,Yan Y H,Xu G Y et al.Toll-like receptor(TLR)-4 mediates anti-β2GPI/β2GPI-induced tissue factor expression in THP-1 cells[J].Clin & Exp Immunol,2011;163(2):189-198.
6 Xu G,Wen H,Zhou H et al.Involvement of IRAKs and TRAFs in anti-β2GPI/β2GPI-induced tissue factor expression in THP-1 cells[J].Thromb Haemost,2011;106(6):1158-1169.
7 陈东东,周 红,解鸿翔 et al.MAPKs途径在anti-β2GPI/β2GPI刺激THP-1细胞表达TF过程中的作用探讨[J].中国免疫学杂志,2012;28(2):99-103.
8 Lopez-pedrera C,Buendra P,Cuadrado M J et al.Antiphospholipid antibodies from patients with the antiphospholipid syndrome induce monocyte tissue factor expression through the simultaneous activation of NF-κB/Rel proteins via the p38 mitogen-activated protein kinase pathway,and of the MEK-1/ERK pathway[J].Arthritis Rheum,2006;54(1):301-311.
9 郭东琳,周 红,周 芳 et al.NF-κB在因子Ⅶa促进SW620细胞增殖迁移中的作用探讨[J].中华肿瘤杂志,2011;33(9):649-653.
10 Raetzsch C F,Brooks N L,Alderman J M et al.Lipopolysaccharide inhibition of glucose production through theToll like receptor-4,myeloid differentiation factor 88,and nuclear factor kappaB pathway[J].Hepatology,2009;50(2):592-600.
11 Lambrianides A,Carroll C J,Pierangeli S S et al.Effects of polyclonal IgG derived from patients with different clinical types of the antiphospholipid syndrome on monocyte signaling pathways[J].J Immunol,2010;184(12):6622-6628.
12 Kristi L Allen,Anne Hamik,Mukesh K et al.Endothelial cell activation by antiphospholipid antibodies is modulated by Kr ppel-like transcription factors[J].Blood,2011;117(23):6383-6391.
猜你喜欢
原子与分子物理学报(2021年2期)2021-03-29 07:30:46
天津医科大学学报(2019年6期)2019-08-13 07:04:42
中成药(2018年7期)2018-08-04 06:04:18
中成药(2018年3期)2018-05-07 13:34:18
中国医疗保险(2017年5期)2017-05-17 08:26:39
三峡大学学报(自然科学版)(2016年6期)2016-04-16 05:02:56
中国康复理论与实践(2015年10期)2015-12-24 05:42:46
安徽医科大学学报(2015年9期)2015-12-16 11:09:42
现代电生理学杂志(2015年1期)2015-07-18 11:02:16
遗传(2014年3期)2014-02-28 20:59:01
