
IFITM3基因的克隆及其在真核细胞中的表达与定位研究①
2012-11-27 11:15:18孙丹丹任静强杜寿文秦艳青王茂鹏田宇飞金宁一
中国免疫学杂志 2012年11期
朱 娜 李 昌 郭 焱 李 沂 孙丹丹 任静强 杜寿文 秦艳青 王茂鹏 田宇飞 金宁一
(军事医学科学院军事兽医研究所,长春130122)
细胞通过模式识别受体(Pattern recognition receptors,PRRs)识别病原微生物的病原相关分子模式(Pathogen associated molecular patterns,PAMPs),诱导信号级联反应,激活转录因子,进而激活特定的细胞因子和趋化因子的表达,使机体处于抗感染状态。其中最主要的就是诱导干扰素(Interferon,IFN),特别是Ⅰ型IFN(IFNα/β)的表达。Ⅰ型IFN通过特异性结合到病毒感染和非感染细胞表面的干扰素受体(Interferon receptor,IFNR),从而诱导数百个干扰素刺激基因(Interferon stimulated genes,ISGs)的表达,启动或调节机体的免疫应答[1]。在ISGs中,干扰素诱导的跨膜蛋白(Interferon-induced transmembrane proteins,IFITMs)是其中较为重要的一种,其表达后除调节细胞的粘附、影响细胞分化外,还与体内细胞信号转导、肿瘤的形成、生殖细胞的归巢和成熟、骨盐的沉积以及抗病毒反应等活动有关[2]。
IFITM3是近期发现的一种具有广谱抗囊膜病毒活性的蛋白。在人体内,IFITM基因家族含有4个成员,即 IFITM1、IFITM2、IFITM3 和 IFITM5[3]。已经证实IFITM1、IFITM2和IFITM3可以由Ⅰ型或Ⅱ型IFN诱导产生,它们对甲型流感病毒的多个亚型(H1、H3、H5及H7)都有很强的抑制作用,其中IFITM3的抑制效果最强,同时IFITM3还对登革热病毒(Dengue Virus)、西尼罗河病毒(West Nile Virus)、SARS-Co冠状病毒(SARS Coronavirus)、人免疫缺陷病毒(HIV-1)等其他不同类型的病毒均有抑制效果[4-6]。缺失IFITM3后IFN的抗病毒反应会明显变弱,所以IFITM3在干扰素反应中发挥着重要作用,为人体抗病毒免疫所必需[7]。
为此,我们通过人Ⅰ型 IFN(rhIFNα2B)诱导293T细胞,提取细胞总RNA,反转录为cDNA,进行IFITM3的扩增,并将其构建到真核表达载体中,开展其表达与鉴定的研究,该结果为进一步研究IFITM3基因的功能和生物学活性以及作用机制奠定了坚实基础,目前国内尚未见相关研究报道。
1 材料与方法
1.1 细胞、菌种和载体 大肠杆菌E.coli DH5α、人胚胎肾细胞293T由本实验室保存;pVAX1载体为Invitrogen公司产品;pMD18-T simple Vector为TaKaRa公司产品。
1.2 主要试剂和仪器 rhIFNα2B干扰素购自Peprotech公司;RIPA裂解液购自Beyotime公司;质粒回收试剂盒购自Axygen公司;质粒制备试剂盒购自北京百泰克生物技术有限公司;胎牛血清(FCS)、胰蛋白酶和DMEM培养基购自Hykolong公司;opti-MEM培养液购自Gibco;FuGEENE HD转染试剂购自Roche公司;辣根过氧化物酶标记的山羊抗鼠IgG、辣根过氧化物酶标记的山羊抗兔IgG和FITC标记山羊抗兔IgG购自中杉金桥生物技术有限公司;兔源IFITM3 PolyClonal Antibody购自武汉三鹰生物技术有限公司;鼠源Anti-beta-actin Monoclonal Antibody购自联科生物;抗荧光猝灭封片液购自碧云天。
英国TECHNE TC512 PCR仪;荧光显微镜为Olympus公司产品;Thermo Scientific公司的NANODROP 2000 Spectrophotometer;激光共聚焦显微镜为德国Leica公司产品。
1.3 IFITM3基因的扩增 人胚胎肾细胞细胞系293T细胞贴壁培养于含10%新生小牛血清及1%青、链霉素的 DMEM 培养基中,于37℃、5%CO2及饱和湿度的细胞培养箱中进行培养。将指数生长期的293T细胞接种于6孔细胞培养板中,每孔接种1×106个细胞,培养过夜。第2天将 rhIFN-α2B稀释液5 μl(浓度1.33×10-5g/ml)加到细胞培养上清液中,孵育12小时。TRIzol法提取细胞RNA,反转录成cDNA,进行PCR扩增,上游引物:ggggatccgccaccatgaatcacactgtccaaac;下游引物:gggaattcctagtggtggtggtggtggtgtccataggcctggaaga。其中上游引物含有EcoRⅠ酶切位点和kozak序列,下游含BamHⅠ酶切位点及his-tag标签,引物由上海生工生物工程技术服务有限公司合成。
PCR反应体系为:cDNA模板 1 μl,2.5 mol dNTP 2.5 μl,rTaq Buffer 2.5 μl,rTaq 1μl,上、下游引物各1 μl,RNase-free 水16 μl,使反应体系终体积为25 μl,离心混匀。PCR程序为:95℃预变性5分钟,94℃变性 30秒,56℃退火 30秒,72℃延伸 45秒,扩增30个循环,72℃后延伸10分钟,4℃保存。将PCR产物与pMD18-T simple Vector载体连接,命名为pMD-IFITM3,转入DH5α感受态细胞,经卡那霉素抗性筛选,挑取阳性单菌落培养,提取质粒,用BamHⅠ和EcoRⅠ酶切初步鉴定正确后,送往北京华大中天生物公司测序。
1.4 重组质粒的构建 用BamHⅠ和EcoRⅠ双酶切经测序正确的pMD-IFITM3,凝胶回收目的片段,并与相同酶切的pVAX1相连,命名为pV-IFITM3,然后转入DH5α感受态细胞,卡那霉素抗性筛选,挑取阳性单菌落培养,提取质粒,并用 BamHⅠ和EcoRⅠ酶切鉴定。
1.5 质粒的制备与纯化 常规方法大量制备重组质粒pV-IFITM3,详细操作步骤参见北京百泰克质粒大量制备试剂盒说明书。然后测定质粒浓度及D260/D280值,-20℃保存备用。
1.6 细胞转染 293T细胞生长至80%~90%融合时,取适量重组质粒pV-IFITM3,利用FuGEENE HD转染试剂转染,质粒和FuGEENE HD比例为3μg质粒和8 μl转染试剂,孵育8小时,设载体pVAX1质粒转染及未转染细胞作为阴性对照。
1.7 IFITM3的RT-PCR检测 采用TRIzol法提取细胞总RNA,RT-PCR扩增IFITM3基因,鉴定基因的转录情况。IFITM3引物及扩增方法参照1.3进行。
1.8 Western blot检测IFITM3在293T细胞中的表达 重组质粒pV-IFITM3转染293T细胞,方法同1.6,转染后培养48小时后,提取细胞蛋白,取2 μl细胞蛋白提取液用分光光度计测定样品蛋白含量,测蛋白浓度 >5 mg/ml。取蛋白样品(600 μg)按4∶1溶于5×loading buffer样品上样缓冲液中,混匀,沸水煮沸10分钟,15%SDS-PAGE凝胶电泳,将分离的蛋白电转移至NC(0.45 μm)膜上,5%脱脂奶粉室温封闭2小时,然后分别与5%脱脂奶粉2 000×稀释的IFITM3抗体4℃孵育12小时,PBST洗涤3次,每次5分钟,加入5%脱脂奶粉2 000×稀释的二抗室温孵育3小时,用PBST漂洗3次,每次10分钟,化学发光ECL法显影检测IFITM3表达,并以β-actin作为内参照。
1.9 重组蛋白的激光共聚集检测 将指数生长的293T细胞接种于6孔细胞培养板中,孔底附有玻片,每孔接种1×106个细胞,培养至细胞生长80% ~90%融合时,进行转染实验,方法同1.6,转染后培养24小时,待细胞生长到接近长满单层时,取出玻片,用 PBS液冲洗 3次,分析纯甲醇(-20℃)固定30分钟,再用 PBS液洗涤3次,用5%脱脂奶粉封闭2小时,PBS液冲洗3次,每次5分钟,加入5%脱脂奶粉2 000×稀释的IFITM3一抗,室温作用2小时后,PBS液冲洗3次,加入5%脱脂奶粉200×稀释的FITC标记羊抗兔IgG和200×稀释的0.05%的伊文氏兰(终浓度0.025%),室温避光作用2小时,冲洗3次,吸干,滴加抗荧光猝灭封片剂,倒置于载玻片上,激光共聚焦显微镜观察、照相。FITC所用激发波长为488 nm,在图像分析系统中选择目标区域取图,并做Z轴方向的光切片合成。
2 结果
2.1 目的基因的获得 琼脂糖凝胶电泳结果显示,rhIFN-α2B处理的293T细胞提取RNA,RT-PCR扩增后,在400 bp处可见明显的扩增条带(图1A),扩增片段的电泳结果与理论值相符,证明得到IFITM3基因。将IFITM3克隆入pMD18-T载体上,经BamHⅠ/EcoRⅠ双酶切,凝胶电泳结果显示,在400 bp和3 100 bp处可见明显条带(图1B),证明IFITM3片段已连接至T载体中。测序结果如图2示,与Gen-Bank中 IFITM3完全一致,证明成功获得 IFITM3基因。
2.2 重组质粒pV-IFITM3的鉴定 质粒pV-IFITM 3经BamHⅠ和EcoRⅠ双酶切后,可见400 bp左右的目的条带(图3),表明构建成功。
2.3 RT-PCR鉴定IFITM3基因在293T细胞中的转录 提取转染重组质粒pV-IFITM3的293T细胞的mRNA,经RT-PCR扩增IFITM3,琼脂糖凝胶电泳结果显示,RT-PCR扩增后在400 bp处有明显条带,但是在空载体pVAX1及阴性细胞对照中也同样出现目的条带(图4A),表明293T细胞含有内源性IFITM3的表达,但与转染IFITM3的质粒相比,条带较弱,表明IFITM3基因在293T细胞中获得转录,设β-actin为内参对照(图4B)。
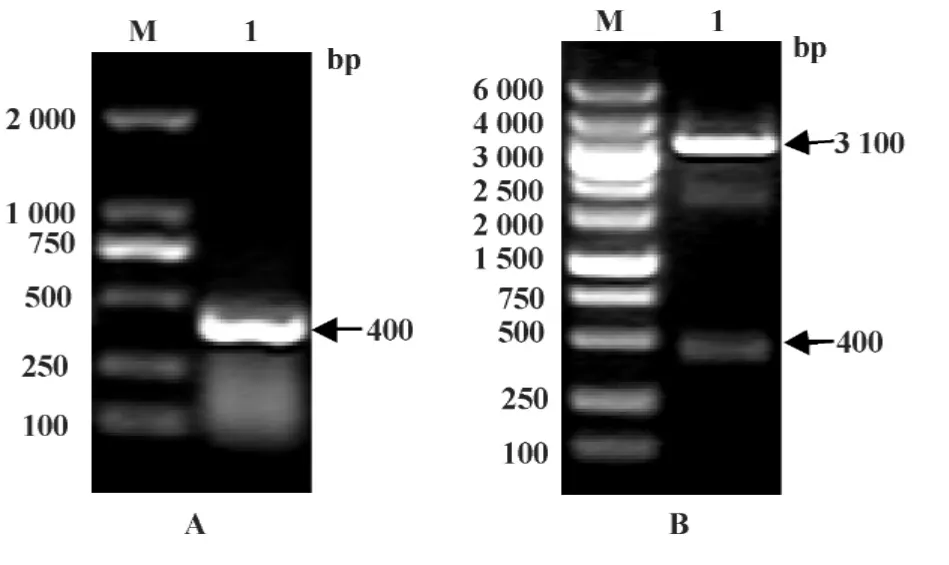
图1 IFITM3扩增及pMD-IFITM3质粒酶切鉴定Fig.1 Amplification of IFITM3 and restriction enzyme digestion of pMD18-T-IFITM3
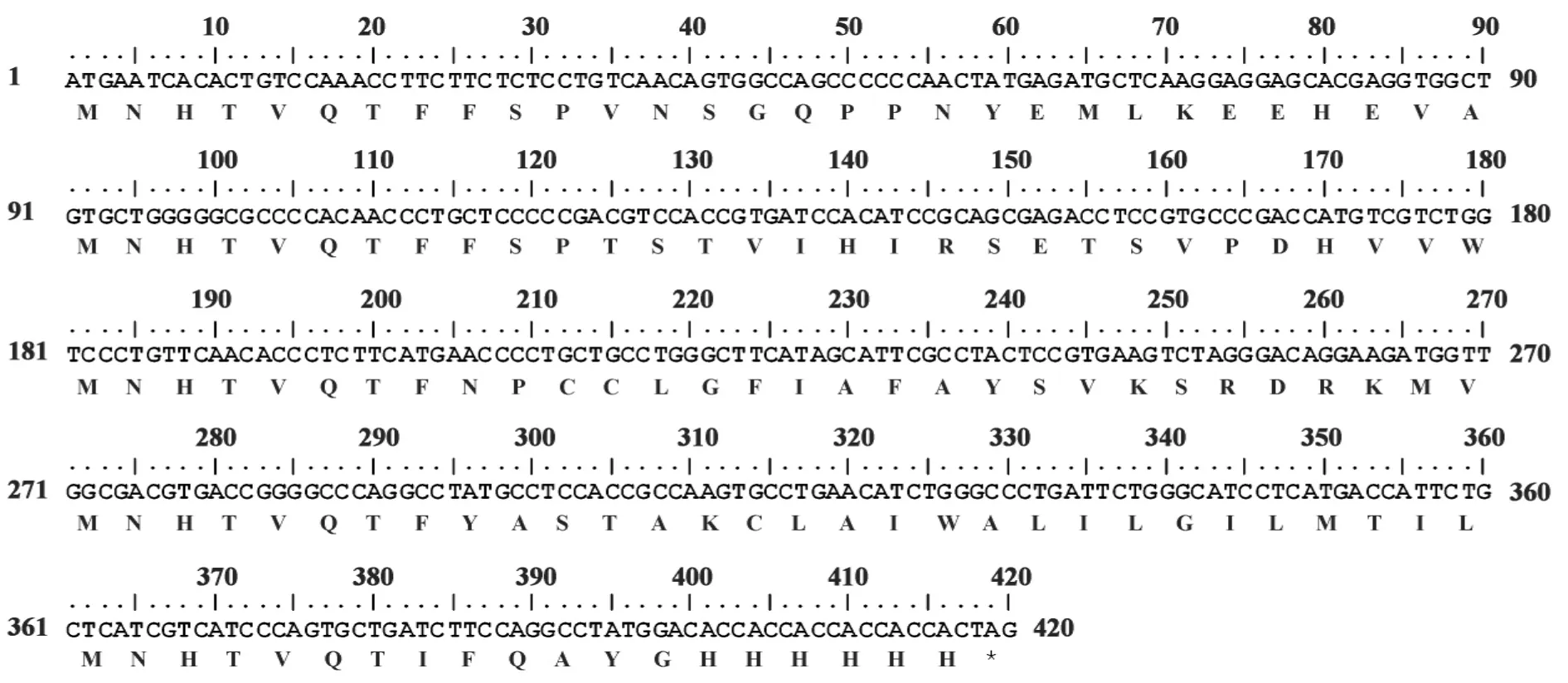
图2 IFITM3的核酸与氨基酸序列Fig.2 Nucleic acid and amino acid sequence of IFITM3
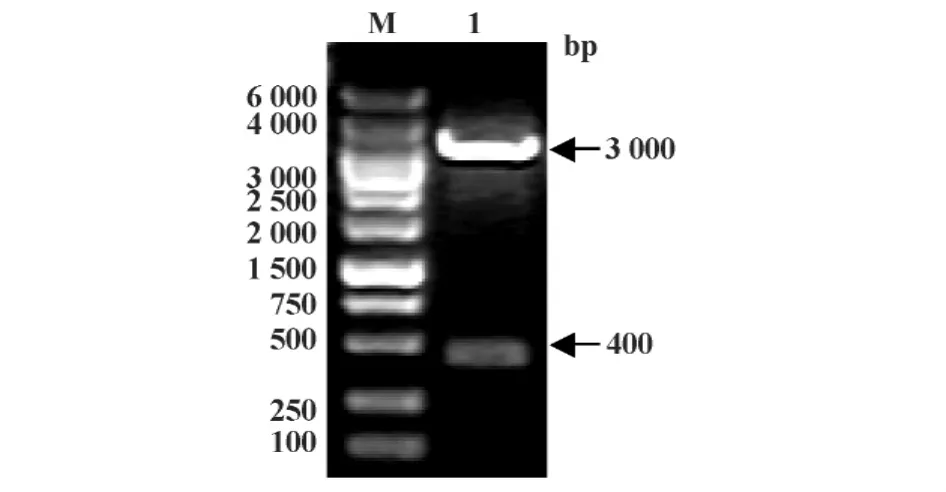
图3 pV-IFITM3质粒酶切琼脂糖凝胶电泳Fig.3 Agarose gel of pV-IFITM3 restriction enzyme digestion
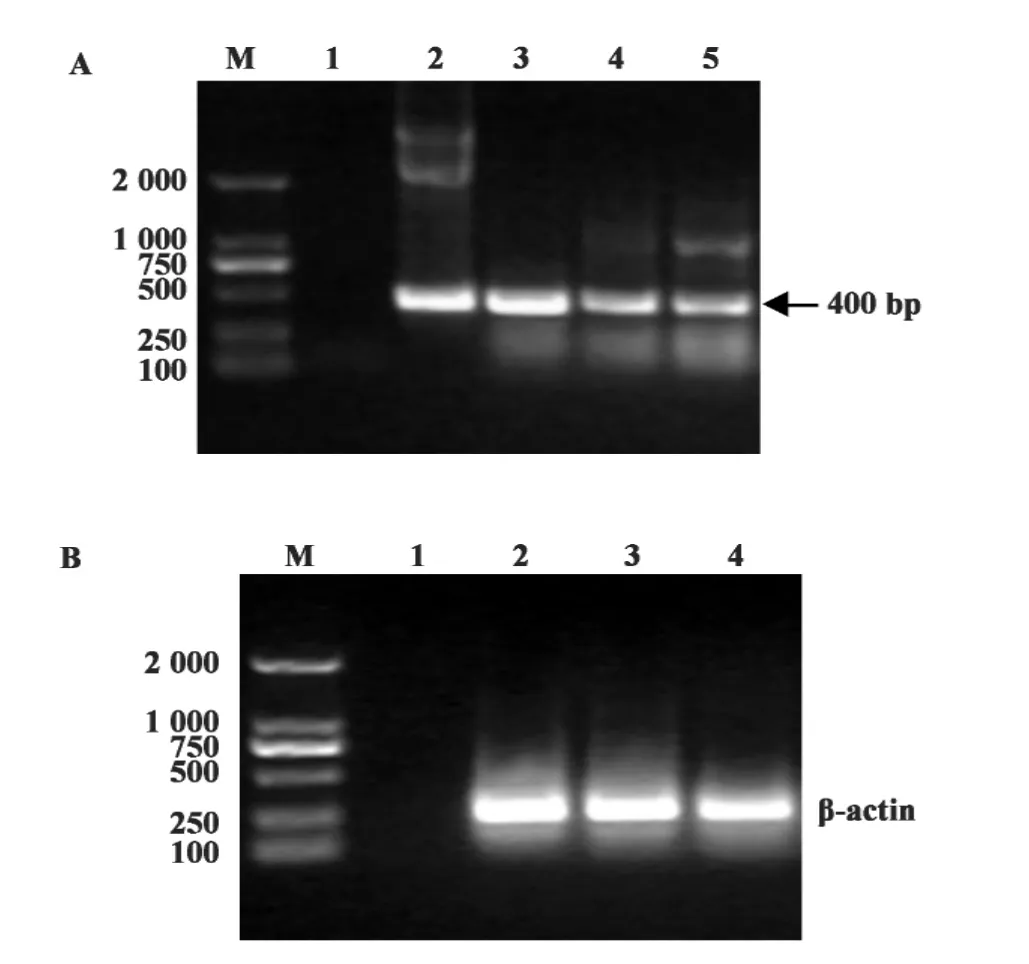
图4 IFITM3基因的mRNA转录检测Fig.4 mRNA transcription identification of IFITM3 gene
2.4 IFITM3基因表达蛋白的免疫印迹检测 利用Western blot对转染pV-IFITM3质粒的293T细胞蛋白进行表达鉴定。化学发光ECL法显影检测结果如图5所示,样品1和2在17 kD有明显条带,表明IFITM3基因在293T细胞中表达,且具有良好的抗体结合能力。另外,该结果与转录水平的检测有所不同,未能检测到内源性IFITM3蛋白水平的表达。表明内源性IFITM3存在mRNA水平的表达,但其由于某种原因不能够翻译成蛋白或翻译后很快被降解,从而在蛋白水平检测不到,但具体原因有待于更进一步研究。
2.5 IFITM3蛋白的定位 利用间接免疫荧光结合激光共聚集对转染pV-IFITM3质粒后293T细胞表达的IFITM3蛋白进行定位研究。由图6可以看出,IFITM3基因转染293T细胞48小时后,表达的外源蛋白主要分布于细胞膜上。

图5 IFITM3基因转染293T细胞48小时后蛋白Western blot检测Fig.5 Western blot analysis of the expression of the 48 h protein of 293T cell transfected by IFITM3 gene
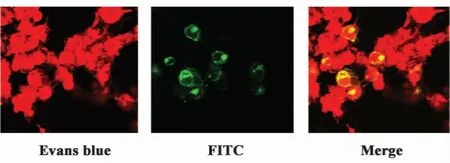
图6 IFITM3基因转染293T细胞48小时后蛋白免疫荧光检测Fig.6 IFA analysis of the expression of the 48 h protein of 293T cell transfected by IFITM3 gene
3 讨论
干扰素(IFN)是一类重要的具有生理功能的细胞因子,其不仅可以调节机体的免疫反应,而且可干扰病毒的复制,被广泛应用于疾病的预防与治疗。但干扰素本身没有直接的抗病毒作用,其主要通过与细胞的干扰素受体(IFNR)结合,从而诱导大量的干扰素刺激因子(ISGs)的表达,通过这些产生的ISGs达到抗病毒的目的。干扰素诱导跨膜蛋白(IFITM)就是ISGs中的一类。在人体中已发现的4个IFITMs中(IFITM1、IFITM2、IFITM3、IFITM5),除 IFITM5在骨盐沉积中的作用研究的比较清楚外,其他三个IFITMs的研究还不够深入[8]。近期研究表明,IFITM3在抑制大多数囊膜病毒早期感染过程中发挥着重要作用,过表达IFITM3能够抑制病毒感染,敲低或敲除IFITM3后,病毒对细胞的敏感性显著增强,进一步研究表明,IFITM3系干扰素发挥抗病毒活性所必需,但IFITM3如何抑制病毒感染却不清楚[9,10]。因此,开展 IFITM3 相关研究,将为更好地理解病毒与宿主之间的关系提供重要线索,为此类分子的抗病毒临床应用提供理论依据。
本研究着眼于IFITM3基因的克隆及其在真核细胞中的表达与定位,利用人干扰素α2B(rhIFN-α2B)诱导体外培养的人胚肾293T细胞,采用RTPCR技术扩增获得干扰素诱导的跨膜蛋白IFITM3全长cDNA,将IFITM3基因亚克隆至真核表达载体pVAX1中获得重组表达质粒pV-IFITM3,转染293T细胞。RT-PCR检测结果表明,IFITM3得到正确转录,同时空载体转染组、未转染组均出现IFITM3 mRNA的表达,但转染组mRNA的表达量明显高于对照组,表明在293T细胞中存在内源性IFITM3在转录水平的表达。进一步蛋白水平的免疫印迹检测结果显示,只有转染含有目的基因质粒(pVAX1-IFITM3)出现目的条带,而对照组无,一方面表明构建的目的基因可以成功表达且具有抗原活性。另一方面也说明虽然静息的293T细胞中存在IFITM3 mRNA的表达,但由于某种原因其不能翻译为蛋白,或者翻译的蛋白量少,或者蛋白翻译后被快速降解,致使在转染48小时后不能检测到,但具体原因有待于进一步研究。该研究提示,在人类正常细胞中即使存在IFITM3内源性mRNA的表达,但由于其不能够进一步大量翻译为具有生物功能的蛋白,从而限制了其抗病毒活性的发挥,所以导入外源性的IFITM3,将有可能改变这一现状,从而为新型抗病毒药物的研制以及转基因抗病毒育种研究提供了重要线索。
在进行IFITM3的定位实验中,选用转染24小时后的细胞蛋白进行FITC抗体标记检测,激光共聚焦显微镜观察显示,IFITM3为遍在表达(结果略),这与之前其他人研究结果相同[4-6],但到48小时后,IFITM3蛋白大都集中于细胞膜上,其机制还需要深入探讨。
总之,本研究在国内率先成功获得了人源的IFITM3基因,并在真核细胞中进行了表达,RT-PCR、Western blot均表明该基因能够表达,且具有良好抗原活性;激光共聚焦显微镜观察,IFITM3基因在后期主要分布于细胞膜上,该研究为基于IFITM3的抗病毒药物研发以及探讨其抗病毒机制研究奠定了坚实基础。
1 Lewin A R,Reid L E,McMahon M et al.Molecular analysis of a human interferon-inducible gene family[J].Eur J Biochem,1991;199(2):417-423.
2 Moffatt P,Gaumond M H,Salois P et al.Brila novel bone-specific modulator of mineralization[J].Bone Miner Res,2008;23(9):1497-1508.
3 Friedman R L,Manly S P,McMahon M et al.Transcriptional and posttranscriptional regulation of interferon induced gene expression in human cells[J].Cell,1984;38(3):745-755.
4 Abraham L B,I-Chueh H,Yair B et al.IFITM proteins mediate the innate immune response to influenza A H1N1 Virus,West Nile Virus and Dengue Virus[J].Cell,2009;139(7):1243-1254.
5 Jennifer L,Qinghua P,Liwei R et al.The IFITM proteins inhibit HIV-1 infection[J].J Virology,2011;85(5):2126-2137.
6 Huang I C,Bailey C C,Weyer J L et al.Distinct patterns of IFITM-mediated restriction of filoviruses,SARS Coronavirus,and influenza A virus[J].PLoS Pathog,2011;7(1):101258-101371.
7 Mith R A,Young J,Weis J J et al.Expression of the mouse fragilis gene products in imnmne cells and association with receptor signaling complexes[J].Genes Immun,2006;7(2):113-121.
8 Andreu P,Colnot S,Godard C et al.Identication of the IFITM family as a new molecular marker in human colorectal tumors[J].Cancer Res,2006;66(4):1949-1955.
9 Fredy S,Martin E,Ulrich C et al.Phylogenetic analysis of interferon inducible transmembrane gene family and functional aspects of IFITM3[J].Cyto,2009;7:314.
10 Suresh C,Kairong C,Keji Z.Transcriptional enhancer factor 1(TEF-1/TEAD1)mediates activation of IFITM3 gene by BRGl[J].FEBS Lett,2008;582(2):391-397.
猜你喜欢
肝博士(2022年3期)2022-06-30 02:48:52
肝博士(2021年1期)2021-03-29 02:32:14
肝博士(2020年4期)2020-09-24 09:21:26
疯狂英语·初中天地(2020年3期)2020-05-21 03:36:52
系统工程与电子技术(2016年2期)2016-04-16 05:16:53
中国继续医学教育(2015年6期)2016-01-07 07:38:50
哈尔滨医药(2015年6期)2015-12-01 03:58:19
中国医疗美容(2015年2期)2015-07-19 10:11:59
中国光学(2015年1期)2015-06-06 18:30:20
河南医学研究(2014年4期)2014-02-27 14:52:28
