
加拿大小麦生产和锈病防治
2012-08-08 06:38AllenXueDawnChi张淑珍李卓夫徐鹏飞姜良宇范素杰
东北农业大学学报 2012年1期
Allen G.Xue,Dawn T.Chi,张淑珍,李卓夫,徐鹏飞,姜良宇,范素杰,王 欣
(1.加东谷物与油料研究中心,加拿大农业与农产食品部,渥太华 安大略 K1A 0C6,加拿大;2.东北农业大学大豆研究所/大豆生物学教育部重点实验室,哈尔滨 150030;3.东北农业大学农学院,哈尔滨 150030)
17世纪,小麦作为一种种植作物的先驱,进入到加拿大的农业生产。小麦是加拿大产业及商业经济的支柱,在国民经济中具有重要地位和作用,它目前是在加拿大所有的10个省份中种植面积最大的作物。在加拿大西部的草原省份,如萨斯喀彻温省、阿尔伯塔省、曼尼托巴省的经济及文化发展中扮演着重要的角色[1]。在加拿大小麦种植面积大约有1 000万hm2,其中700万hm2六倍体春小麦,200万hm2硬小麦,100万hm2冬小麦。在2010年,这些小麦的产值大约有80亿[2-3]。由于小麦巨大的种植面积及其在加拿大经济中的重要性,关于小麦品种的选育已得到产业及政府科学家的高度重视。种植者通过西方谷物研究基金会每年对小麦育种提供超过300万美元的资助[4]。大部分种植计划的目标包括:①继续防治新的锈病小种;②选育抗病性强的新品种;③在产量和质量上不断提高。
加拿大小麦加工成面粉、谷类食品、饲料、面包、意大利面和烘焙产品,在加拿大及全球70多个国家销售。硬质小麦用来做通心粉和粗面条,红色的优质硬质小麦用于做高质量的面包,而草原春麦做饲料小麦。商业出口的小麦只产于两个主要的区域,即草原区,由曼尼托巴、萨斯喀彻温以及英属哥伦比亚东北部以及在加东,主要是安大略的南部。不同于加拿大的其他作物,联邦法律规定,加拿大西部的小麦生产商进行市场销售必须通过加拿大小麦委员会。加拿大小麦委员会是用来监测由西部农民种植的所有的小麦和大麦市场的组织。加拿大是世界上第三大小麦出口国,占全球小麦出口量的20%[5]。
1 小麦的分布和生产情况
草原省的小麦产量占加拿大小麦生产总量的90%以上[6-7]。大约85 000户农民每年在草原省生产2 200~2 400万t小麦。在大草原主要种植春小麦和普通小麦。在小麦的南部干旱生产区的萨斯喀彻温省、阿尔伯塔省,大量的硬质粗粒小麦被制成通心粉。在草原三省中,萨斯喀彻温省小麦的产量最高(6 500 000 hm2或46%),其次是阿尔伯塔省(2 500 000 hm2或27%)和曼尼托巴省(1 500 000 hm2或18%)具体见图1[7]。
在加拿大东部,种植的多为秋季播种的冬麦,主要种植在安大略的南部和中部(360 000 hm2或8%),在安大略东部,魁北克及曼尼托巴省份(50 000 hm2或1%)春季播种的小麦也有一定的数量[8]。

图1 1908~2007年加拿大各地区小麦产量Fig.1 Wheat production in Canada 1908-2007
小麦可以种植在各种土壤类型,但最适宜种植在未曾播种过作物、排水良好的土壤,小麦因是轮作体系的重要作物,所以其播种面积波动不大。作物种植面积小范围的波动是组成作物轮作的重要部分。除此之外,在萨斯喀彻温的某些年份干旱成为产量的限制因素[6-7]。天气条件是引起产量变化的原因,特别是萨斯喀彻温省和西部的大草原频繁的干旱,曼尼托巴省南部的洪水。虫害问题不断出现,通过选育抗虫品种是很重要的,锈病不断改变并适应性逐渐增强[9-10]。新病害例如由镰刀菌引起的赤霉病在小麦的主产区也不断发生[11]。抗除草剂的杂草也成为一个问题。最近,利用生物技术作为一种育种工具已备受争议,携带草甘膦基因的小麦得到消费者的关注[12]。
2 小麦品种类别和代表性品种
自从100年前,小麦在加拿大种植,六倍体面包春小麦就占有主要的市场,是小麦的优势品种,在整个时期都很明显,尽管硬粒小麦在此时期种植面积显著增加以及六倍体冬小麦也占有小麦面积的相对一小部分,而冬小麦只占种植面积很小的一部分。根据不同的质量标准和多样化的市场需要,六倍体小麦已经被划分成很多类,包括加西硬红春小麦、加平原春红小麦、加平原春白小麦、加西超强筋小麦、加西软白春小麦、加西硬白春小麦、加西杜伦小麦和加西红冬小麦[13-14]。加西硬红春小麦和加西杜伦小麦是目前的主要品种,占加拿大西部小麦种植面积的88.3%(见图2)[14]。剩余的6个种类品种只占11.7%,降低到与特用品种相同的地位。
2.1 加拿大西部红春小麦(Canada western red spring,CWRS)
加拿大西部红春小麦品种作为第一个小麦品种于1813年在加拿大西部地区种植[15]。加拿大西部红春小麦适种地区较为广泛,具有较好的面粉加工特性,适用于许多不同工艺条件下各种烧烤类食品的生产。因此,CWRS小麦在世界贸易中的价格较高,其种植主要在加拿大的三个草原诸省,并以CWRS栽培品种为主,在1941~2007年期间,平均种植面积为880万hm2,其中1967年最高为1 140万hm2。一些品种广泛种植很长时间,比如,CWRS小麦品种Thatcher是1939~1968年的主栽品种,Neepawa是1970~1990年的主栽品种,Katepwa是1985~1995年的主栽品种,AC Barrie是1997~2007年的主栽品种,Lillian是自2007年以来的主栽品种(见图3)[7]。其他具有代表性的CWRS品种有 Selkirk、Marquis、Red Bobs、Manitou、Rescue、Canthatch、Columbus、Aura、CDC Teal和Superb。
2.2 加拿大平原红春小麦(Canada prairie spring red,CPSR)和加拿大平原白春小麦(Canada prairie spring white,CPSW)
第一个加拿大平原红春小麦品种和加拿大平原白春小麦品种的种植分别为:1985年CPSR小麦品种HY320[16],1988年CPSW小麦品种Genesis[17]。CPS小麦产量高出CWRS小麦产量的20%~30%。1985~1989年,1987~1990年主栽的CPSR小麦品种是 HY320,1990~1993年是 Biggar,1994~2003年是 AC Taber,1998~2006 年是 AC Foremost,2000~2005年是AC Crystal,2006~至今是5700 PR。1991~1998年普遍种植的CPSW小麦品种是Genesis,1998~2002年是AC Karma。CPSW小麦品种中多数栽培品种与HY320相关[7]。1985~2007年已播种CPSR和CPSW栽培品种的地区平均面积为500 000 hm2,1997年最高为900 000 hm2[7]。目前,高产的CPSR和CPSW品种被评估作为生产酒精的饲料原料[18]。
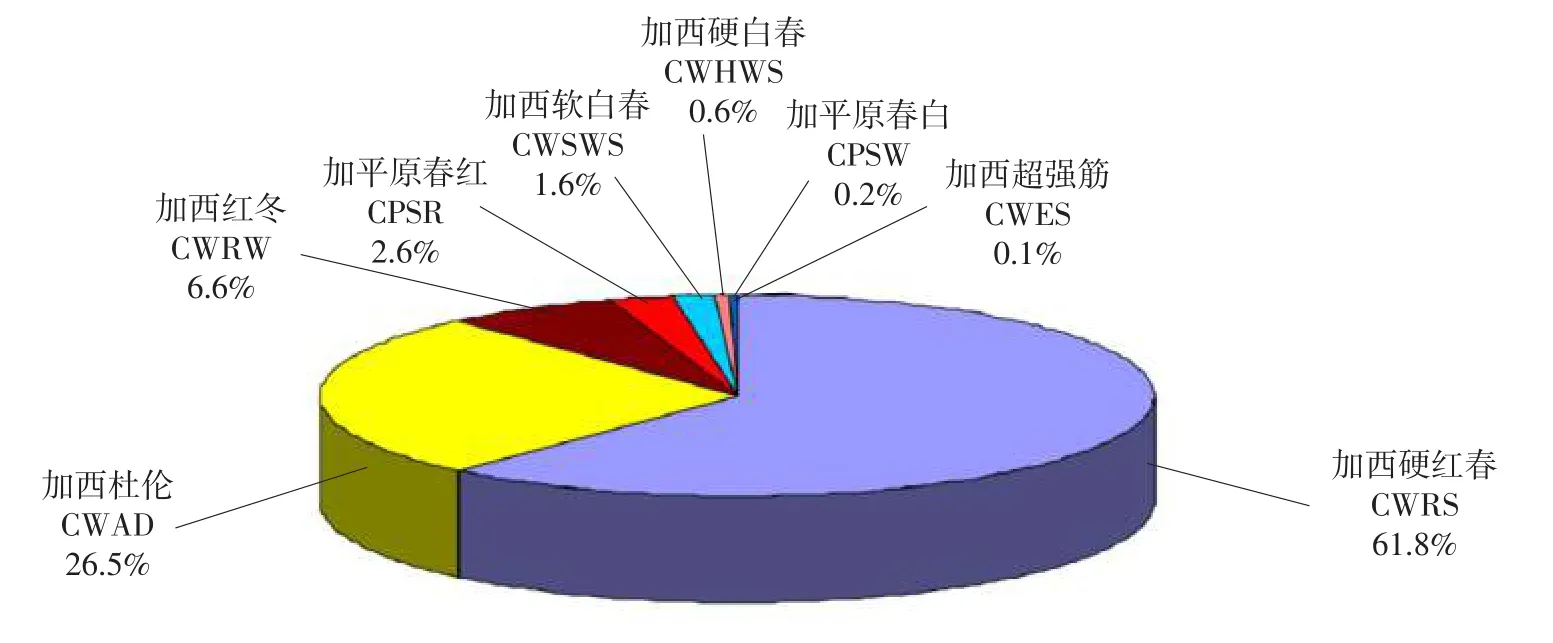
图2 加拿大小麦市场划分Fig.2 Canadian wheat market classes
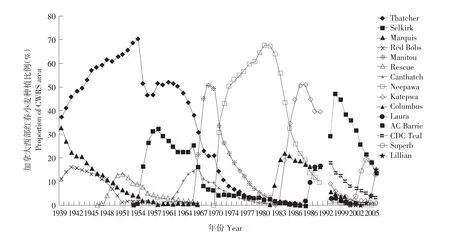
图3 1908~2007年加拿大西部红春小麦(CWRS)种植比例Fig.3 Canada western red spring wheat(CWRS)production in Canada 1908-2007
2.3 加拿大西部超强筋小麦(Canada western extra strong,CWES)
CWES小麦品种面筋弹性较强,中等蛋白含量,使它不同于CWRS的面包制作特性。CWES品种开始种植于1993年[19],第一个CWES品种Glenlea注册于1972年。Glenlea具有很好的强筋特性,有利于冷冻生面团的生产,冷冻面团的生产需要强筋性经历冷冻和解冻的循环,其与弱筋性小麦混合后可制作出较满意的面包类产品[20]。1998~2002年,2005~2006年,Glenlea是主要的CWES栽培品种。2003~2004年,CWES的主栽品种是Bluesky,2007年CWES的主栽品种是CDCRama。在播种CWES品种的地区,该品种种植面积呈下滑趋势,从1995~2000年平均每年250 000 hm2,到2001~2007年平均每年100 000 hm2[7]。这一现象说明,由于其他国家强筋性品种生产和北美科学技术进步等原因,导致对于冷冻生面团生产所需的强筋性面筋的需求有所降低。
2.4 加拿大西部软白小麦(Canadian western soft white spring,CWSWS)
加拿大西部软白小麦几乎只在阿尔伯塔省的南部灌溉条件下种植。19世纪30年代Ellison的面粉厂为了满足当地对意大利粉的需求,引进了软白小麦[21]。与其他小麦品种相比,软白小麦的种植面积较小。19世纪70年代末到1996年,CWSWS的主栽品种是Fielder,之后被AC Reed取代。AC Reed作为主栽品种,在2003年被AC Andrew取代。其他产量和品质较好的软白小麦品种有AC Phil,AC Meena,AC Nanda和AC Bhishaj。近年来,在种植软白小麦地区,该品种的种植面积有所减少,从1979~1999年平均种植面积100 000 hm2,降低到2000~2005年的20 000 hm2,这主要是因为加拿大西部红春小麦和加拿大西部杜伦硬质小麦(Canada western amber durum,CWAD)的发展和广泛种植。CWRS和CWAD可以在灌溉条件下种植,并且售价较高。目前,CWSWS主要用于生物燃料乙醇的生产,特别是AC Andrew品种,它具有高产和低蛋白含量特性[18]。
2.5 加拿大西部硬质白皮春小麦(Canada western hard white spring,CWHWS)
2001年开始种植加拿大西部硬质白皮春小麦品种,以Kanata[22]和Snowbird[23]品种的审定为标志,其目标市场为生产亚洲风格的面粉条和面包类食品。加拿大西部硬质白皮春小麦的种植面积从2001年的20 000 hm2增长至2005年的330 000 hm2。其中95%以上面积播种Snowbird品种(2003~2007年为98%)。加拿大西部硬质白皮春小麦的面包烘焙品质与CWRS相似,不过种皮为白色。这一小麦品种的未来取决于对硬质白皮小麦的需求以及草原生产者对其的接受程度。
2.6 加拿大西部杜伦硬质小麦(Canada western amber durum,CWAD)
在加拿大,小麦种植初期,硬质小麦就种植于加拿大西部地区。从20世纪初至20世纪60年代广泛种植的硬质小麦从多个地区引进,包括Kubanka、Arnautka(俄罗斯)、Mindum(美国)、Pelissier(阿尔及利亚)、Golden Ball(南非)、 Carleton(美国)、Stewart(美国)和Ramsey(美国),在1900年至20世纪60年代期间种植较为广泛[24]。1963年,Stewart 63成为加拿大第一个审定的硬质小麦品种,并于1967年作为硬质小麦的主栽品种广泛种植。20世纪60年代至70年代,Stewart 63和Ramsey是主栽品种,其中Stewart 63品种占草原地区硬质小麦种植面积的68%,Ramsey占13%。1972~1973年Hercules成为主栽品种。1974~1987年Wascana和Wakooma成为主栽品种,其中1976~1986年种植面积达70%。1988~2004年Kyle成为主栽品种,2005年被Avonlea取代。2007年CWAD小麦的主要种植品种为Kyle,AC Avonlea、AC Strongfield和AC Navigator。
2.7 加拿大西部红冬小麦(Canada western red winter,CWRW)
加拿大冬小麦很早就开始种植了,不过仅限于阿尔伯塔省南部地区。曼尼托巴省和安大略省南部地区,这些地区的冬天积雪保护可能性较大。1977年Norstar品种的审定通过,使在生产上超越北美品种的耐寒品种方向迈出了重要一步[25-26]。Norstar很快成为冬小麦的主栽品种,不久之后便大量种植,取代了所有以前的种植品种。1985~1992年,Norstar每年的种植面积占冬小麦种植面积的90%,其他主要的冬小麦品种有1993~2002年的CDC Kestrel,1998~2004 年的 CDC Clair,2002~2004 年和2006~2007年的CDC Falcon,2004~2006年的AC Bellatrix,2007至今的CDC Buteo。CWRW小麦的种植面积的增长较为显著,2004~2007年草原地区平均种植面积500 000 hm2,安大略湖地区种植面积400 000 hm2[7]。近年来,冬小麦的种植面积逐年增长,原因主要是抗病性和品质性状的有效改良,以及乙醇生产对小麦的大量需求。
3 小麦锈病
影响加拿大小麦的锈病主要有三种类型:秆锈病、叶锈病和条锈病[27]。秆锈病的主要症状有:在易感病植株上,茎部形成砖红色孢子堆,叶片小范围染病(见图4a)。叶锈病的病斑仅限于叶片内部较大范围;病斑可能形成于叶鞘部位,但不会感染茎部组织。形成的孢子堆为圆形,比秆锈病孢子堆小,呈现淡黄红色,并在高感部位紧密排列,整个植株叶片呈现橙红色(见图4b)。条锈病就是因为其在小麦叶片及穗部形成长的黄色的孢子堆条纹状孢子堆的特性命名的,其症状主要是在叶片正面形成伸长的黄色孢子堆,并延伸至整个叶片(见图4c)。自加拿大种植小麦以来,锈病就成为每年小麦生产的主要病害之一[9]。病害严重年间,由锈病造成的损失高达70%[28]。
秆锈病主要是由Puccinia graminis f.sp.tritici引起的,叶锈病主要是由P.triticina引起的,条锈病主要是由P.striiformis f.sp.tritici引起的。每一个引起病害的病原菌都以许多小种的形式存在,它们具有不同的致病性,侵染和毒力特性[29]。由于同一锈病种类之间不同生理小种间发生突变和重组,可以导致新的生理小种的产生。如果环境条件发生改变,现存的生理小种传播较快,从而具有更强的毒力,因此,抗性基因可能会失去其抗病性。锈菌属于专性寄生菌,只能在自然条件下在寄主组织中生长和繁殖,它们不能像腐生菌那样生存。在缺少寄主的条件下,病原菌只能以孢子的形式生存很短的时间。

图4 小麦锈病发病症状Fig.4 Symptoms of wheat rusts
3.1 秆锈病
秆锈病是历史上影响加拿大小麦生产的主要问题,曾在1954年,发生了历史上最严重的一次疫情[28]。随着抗性品种的育成,秆锈病不再是毁灭性的病害[10]。在加拿大,Thatcher是第一个具有重要意义的抗秆锈病品种,从1939年到20世纪70年代被广泛种植[7]。多年以来,加拿大的小麦品种对加拿大出现的毒力小种表现了良好的抗性,但随着新的毒力小种不断出现,现有抗性品种失去作用,秆锈病可能会大规模爆发。例如,Ug-99被认为是50年里最为严重的小麦秆锈病毒力类型,自1999年在乌干达鉴定出以后,现已经传播到东非,并向印度次大陆蔓延[30]。最近,这个生理小种已在伊朗发现并且威胁到中东和中亚的小麦生产。Ug-99对现在美国和加拿大种植的大部分小麦品种也是有致病性的[31]。通过气流传播或者人类的活动,Ug-99可能不久会进一步向美国和加拿大迁移。Ug-99对于加拿大的小麦产量是一个潜在的威胁,因而,也影响到世界粮食的供应。
禾柄锈菌,属担子菌亚门真菌,共产生5种类型的孢子:担孢子、性孢子、锈孢子、夏孢子和冬孢子,还需要两个不相关的寄主植物,即小麦和小蘗,来完成它的世代循环[29]。小麦秆锈病的循环是从新小麦上的病原孢子裸露出来开始的。在北美州,小蘗是小麦秆锈病流行的最主要的菌源植物。如果小蘗靠近小麦地种植,它在春天将成为锈孢子的持续来源给小麦带来最初的感染。
在加拿大,即使没有小糵,小麦秆锈病也可以流行[32]。在没有小糵的条件下,美国和墨西哥南部冬小麦上的夏孢子可通过气流传播到加拿大的小麦上,因此春天里小麦仍有病原孢子的存在。墨西哥海湾温和的气候有利于秆锈病病原的存活,并且有助于传播到冬小麦的生产地。春天盛行的南部季风携带着夏孢子到加拿大的草原上,美国大平原上,安大略南部,俄亥俄山谷地区,加拿大东部的州和美国东北部。
小麦秆锈病流行需要白天温度高(25~30℃),夜晚温度适宜(15~20℃),以及有雨水或露水的湿的叶片[33-34]。锈孢子和夏孢子像其他的孢子一样需要液态水才可以萌发,并通过气孔感染。在适宜条件下侵染7~15 d,夏孢子才能产生,所以在一个生长季节可以产生多个世代的接种体。
秆锈病通过几种方式导致小麦减产[35]。病原菌可以从用于籽粒发育的植物细胞中吸收营养。当孢子堆突破表皮组织后,它会影响植物调整它的呼吸作用,从而降低新陈代谢的效率,阻碍输导组织从而使籽粒干瘪。秆锈病也能使小麦的茎秆变弱,使其在强风或大雨中倒伏,严重的倒伏使机械收获困难或难以收获。
3.2 叶锈病
叶锈病在加拿大的小麦种植区每年也有发生,是草原东部(曼尼托巴省和萨斯喀彻温省)和安大略省南部发生最为严重的小麦病害。每年叶锈病都会造成小麦减产1%~20%。硬质小麦是传统的抗叶锈病的抗性品种,而面包麦普遍为感病品种。2001年,在墨西哥的西北部检测出一个对硬质小麦有高致病性的新的叶锈病生理小种[36]。在世界范围内,它是唯一一个对存有许多抗病基因的六倍体小麦没有致病性,却对世界范围收集的大部分硬质小麦品种有很强致病性的生理小种。
叶锈病的病原产生和禾柄锈菌一样的5种孢子,也需要可选择的寄主植物。例如,利用唐松草属(Thalictrum)植物来完成它的周期循环[29]。由于在美国和墨西哥南部夏孢子能在小麦上越冬,叶锈病在加拿大持续的发生[32]。病菌每年春天都会向北方蔓延,并在小麦的成熟过程中发生夏孢子循环。
在加拿大西部,生长季节的6月叶锈病开始侵染,8月到达顶峰。在加拿大,冬孢子会在秸秆上越冬,在春天萌发。每个冬孢子会产生4个担孢子。担孢子不会感染小麦,但是能影响寄主唐松草属植物。唐松草属植物产生孢子后,侵染小麦产生夏孢子。北美本土的唐松草属植物对叶锈病有很高的抵御作用,并能抑制其传播。
在最适温度为10~30℃条件下,叶锈病发展迅速[33-34]。叶锈菌能和小麦叶片在同样的环境条件下存活,如果只是侵染但不产生孢子。在20℃左右的潮湿条件下,叶锈病菌可以在3 h或更少时间内进行侵染,然而在更长期的潮湿条件下病原菌侵染更频繁。在低温下,更长时期的潮湿对于病菌的侵染是必要的,例如10℃,需要12 h的潮湿条件。露期温度在32℃以上或2℃以下几乎没有病原菌的侵染[37]。在15~25℃,夏孢子接触自由水后30 min开始萌发。在适宜及持续温度下,7~10 d内孢子可萌发产孢。10或15℃低温或更长的时间是必须的。当温度临近或低于结冰温度时,病原菌可以以菌丝体的形式存活1个月或更长时间。
小花聚的减少以及籽粒的干瘪是叶锈病导致小麦减产的主要原因。在高感病品种中,小花、分蘖以及植物自身在早期病害流行被杀死。在加拿大,根据作物生长阶段的感染时期和小麦的感病性推算,叶锈病可减产5%~25%[38-39]。
3.3 条锈病
条锈病是低温环境下发生的主要疾病,它是生长在阿尔伯塔和英属哥伦比亚地区的较高海拔处小麦的常见疾病。许多条锈病的生理小种已在加拿大鉴定出来[40]。最近在阿尔伯塔中部和安大略省南部条锈病广泛流行。条锈病也会在萨斯喀彻温省西部发生,偶尔也会在北美东部大草原发病,造成这些地方的小麦减产。这可能预示着新的表现型开始出现,并且更适应加拿大南部温和的环境。直到2010年,条锈菌一直被误认为没有转生寄主。但最近研究发现[41],条锈菌也有转生寄主,包括小糵属的几个种。例如Berberis chinensis,B.bolstii,B.koreana 和 B.vulgaris,这些小糵属的转生寄主被认为对加拿大部分地区及世界上条锈菌新小种的产生起了重要作用。在美国,病原菌以夏孢子或休眠菌丝的形式在当地草地的叶子和冬小麦上过冬。在气候温和的冬季,病原菌能在加拿大大部分地区的冬小麦上过冬。在大多数夏季,夏孢子能通过气流传播到美国的北部以及加拿大。
凉爽的夜晚有利于病原菌的侵染和产孢[29],条锈病的最适温度为9~12℃,高于20℃抑制孢子的形成。孢子需要在潮湿的叶子上几个小时以便萌发,之后感染寄主。在侵染之后,来自于孢子的菌丝沿着与叶脉平行的方向伸长,使叶片出现黄色条纹,然后慢慢扩散直至整个叶片。在适宜的条件下感染7 d后,症状开始出现,8~14 d菌丝大量生长。在黄色的条斑中含有大量的夏孢子,这些夏孢子还可以进一步侵染作物。在生长季节,真菌可以产生几代夏孢子,这些孢子可以继续侵染绿色组织或植物。在加拿大草原和安大略南部,夏天的高温通常能抑制疾病发展以及防止疫情的爆发[32]。除了接种物和感病品种外,温和的冬季和凉爽、潮湿的夏季也是加拿大西部和安大略南部条锈病大爆发的主要原因。
严重的流行疾病导致小麦大量减产,最高可达75%[42]。因为叶片表皮受到破坏,进行光合作用的叶片和蒸腾作用的水大量损失,所以粮食产量大幅降低。
4 锈病的防治策略
由于小麦锈病通过空气传播,隔离病原菌仅仅是延缓,并不能防止病原菌的进入。全球小麦锈病菌的毒力类型、攻击性、适应性存在显著差异。人们应该注意不要在不知情的情况下传播或者让锈病孢子从流行病领域传播出来。种植抗锈病品种是防治锈病的有效方式,但是不要在同一地点持续种植一种抗性品种[43]。也可利用早播种,消除选择寄主和应用杀菌剂等其他的措施来防治该病。这些策略相结合或者整合已有的抗病防治方法和应用杀菌剂将会成为防治小麦锈病最有效的方法。
4.1 抗性育种
在加拿大,小麦育种主要就是提高其抗病能力,尤其是抗锈病。超过一个世纪的时间里,加拿大已经意识到,通过抗性品种的协作来防治锈病是有效的。1916年小麦锈病在加拿大严重发生,1925年主要的锈病实验室在温伯尼建立,之后对于抗锈病的育种不断加强,直到现在[44]。起初,抗病育种主要集中在抗秆锈病但随后抗叶锈病的品种也育成了。
Thatcher是第一个具有重要意义的抗秆锈病品种,从1939年到20世纪70年代被广泛种植。除了20世纪50年代15B的流行和20世纪80年代由于感病品种Norstar引起的冬小麦的减产,一般而言,抗病品种对秆锈病是耐病的,并能长时间抗病,Thatche复杂的遗传抗性形成了对加拿大西部红春麦抗性的基础,这个已经被增加的抗秆锈病基因补充了,例如Sr6、Sr7a、Sr9b、Srll等。1979年,抗秆锈病的加拿大西部的小麦品种每年的价值在217百万美元[45]。现在的每年的抗性价值可能会更高。
叶锈病的抗性基因对于叶锈病有重要的防治作用,但这些抗性基因不像秆锈病的耐病性持久。Lrl、Lr10、 Lr13、Lr14a、Lr16和Lr34是加拿大品种中普遍存在的抗性基因[27]。叶锈病特有的抗性基因Lr21和Lr22a,被整合到加拿大西部红春麦品种。这些基因在20世纪70年代被加拿大学者从节节麦中分离出来[46],而且至今仍对加拿大的叶锈菌也有很高的抗性。自1994年首次在AC Cora品种中鉴别出 Lr21 以来[47],在 McKenzie,Lovitt[48]和CDC Alsask中也相继发现。1991年,在AC Minto首次鉴别出Lr21a后,已广泛应用在5500HR和5600HR中[49],通过抗性的聚合基因图谱鉴定从而可以实现持久的叶锈病的抗性。例如,1991年,注册的含有5个抗叶锈病基因(Lrll、Lr13、Lr14b、Lr30、Lr34)[50]的Pasqua品种。在过去的时间里,对抗性基因Lrl、Lr10、Lr13、Lrl4a、Lr16的毒力不断发展;然而通过对Lr21、Lr22a、Lr34和许多不同抗性基因的聚合在对大多数品种仍有很高的持 久 抗 性[27]。Lr52=LrW[51]、LrW2、Lr18、Lr35 和Lr46会是未来有效的抗病基因。
尽管Yr18基因使许多品种具有部分抗性,但对加拿大小麦品种来说抗条锈病的遗传基础仍是未知的[52]。如果小麦条锈病开始在加拿大大多数产区危害严重的话,那么提高条锈病抗性育种工作将是必要的。
总的来说,通过种植聚合有两个或两个以上抗病基因的抗性品种是防治小麦锈病的有效手段[53]。但育成对新生理小种的抗性品种以及与耐病资源相结合仍是艰巨的任务,虽然加拿大的小麦品种对秆锈病的耐病性已经有50年了,仍需要对秆锈病抗性品种或抗性基因遗传基础进行改善[54]。最近,随着Ug-99生理小种引起的小麦秆锈病在东非大规模爆发,提供了多抗性对于防治该病是有重要意义的一个典型例子[55]。Ug-99对许多Sr基因以及加拿大小麦品种有很高的致病性,一旦引进加拿大将会成为引发秆锈病流行的潜在威胁。因此需要与其他作物有效的抗病基因相结合来拓宽加拿大小麦品种遗传图谱的多样性[56]。
4.2 消灭转生寄主
破坏寄主是控制加拿大秆锈病最重要的方法,如小糵[32]。小糵作为可替代的锈病有两点原因。首先,它作为冬小麦早期的接种源,能诱发更早,更具破坏力的病害流行。其次,由于病原体可以完成有性循环,致使新的生理小种出现。这些生理小种可以侵染一些抗病品种。通过破坏这些寄主可以阻止其完成有性循环和一些新生理小种的演变。
如果没有有性循环,秆锈病菌的最初春天接种源来自遥远的地方,从而推迟在生长季节的流行,如果发生,也会是一个较低水平。小糵是从欧洲引进后被广泛种植在加拿大东部的作物,1918年在加拿大西部和密西西比河州以及20世纪60年代在安大略都开始了根除小糵的行动。加拿大食品检验机构严格控制进口和国内出售小糵,只有对病原体有免疫功能的物种才能在加拿大出售。
4.3 化学防治
在一些病害严重,而小麦生产潜力也较高的区域,主要通过喷洒杀菌剂来防治条锈病。过去的十年里,由于小麦赤霉病的严重流行和商品价格的提高,加拿大的部分地区杀菌剂的用量在不断增加。包含丙环唑和戊唑醇的杀菌剂是比较有效的。然而,随着抗药性的提高,杀菌剂会成为额外的生产成本。
4.4 早期播种
加拿大流行的小麦锈病菌可以在美国南部越冬。到了春天,美国南部的谷类作物开始生长,锈病的病原体和夏孢子被向北吹到美国的中西部,然后到了北部[33]。从美国吹向加拿大的夏孢子不断积累,感染加剧,6月到达顶峰[39],7月末到8月初锈病便在加拿大草原开始大规模流行。如果春天谷类作物提前播种,到了生长季节,接种密度较低,便可避免严重的锈病感染。
5 结论与展望
小麦是加拿大种植面积最大的作物,主要种植在加拿大西部的曼尼托巴、萨斯克彻温、亚伯达等草原省。从加拿大西部种植小麦以来,已有100多年的历史,尽管在这期间硬质麦种植面积显著增加,以及六倍体冬麦也在小麦种植区域占有相对一小部分比例,但六倍体的春面包麦在市场仍占主导地位。根据不同的用途质量评价标准,六倍体小麦的多样性已被划分为数个市场种类。主要的面包春麦是加拿大西部红春麦的一类。一些品种已经种植很长时间了,例如1939~1968年主要种植加拿大西部红春麦Thatcher;1988~2004年主要种植CWAD品种的Kyle。还有多年来特定种植的小麦品种如CWES的Glenlea和CWRW的Norstar。老品种代替新品种的主要原因是产量方面的提高及锈病抗性以及食用质量等的改善。
在加拿大,小麦锈病的遗传抗性对这类疾病有很好的防治作用。尽管在防治锈病方面已取得了很大的进步,要想持续控制小麦叶锈病仍是艰巨的任务。这主要是由于病原菌群体的不断改变而使品种的抗性失去作用。将来其他锈病例如小麦条锈病或者对小麦有高致病性的禾柄锈菌Ug-99生理小种可能给加拿大谷类作物带来新的威胁。长期的研究以及抗性育种是加拿大未来防治锈病最有效的策略。通过利用耐病品种,有效抗性基因的布置以及聚合抗病基因,将是控制该病最为有效的措施。
新的小麦秆锈病、叶锈病、条锈病生理小种出现后,可以穿过陆地、海洋几千米的距离到达加拿大。外源锈病小种的引入会给加拿大小麦的生产和改良带来可怕的影响。新的生理小种可能会对在目前小麦品种或育成品系中不具有的抗性基因具有致病性,使它们在应用于抗性育种之前失去作用。外源生理小种也会使正在生产中种植的小麦抗性基因失去作用,从而缩短一个品种有效的抗病时间。在未来,通过监测世界范围内的小麦生理小种来应对新生理小种给小麦生产带来的潜在威胁是非常必要的。
*本文为特邀加拿大专家综述。
[1] Rawlinson H G,Granatstein J L.The Canadian one hundred:The 100 most influential Canadians of the twentieth century[M].Toronto:McArthur&Company,1997:15-18.
[2] Statistics Canada.Field crop reporting series-3401.Table 001-0010[R].Statistics Canada,2010.
[3] Statistics Canada.2011.Field crop reporting series-Stocks of principal field crops at December 31,2010.Catalogue no.22-002-X,vol.90,no.1.
[4] Western Grains Research Foundation.Western grains research foundation endowment fund policy[EB/OL].[2010-09-30].http://www.westerngrains.com.
[5] Canadian Wheat Board.An open market for CWB grain-A study to determine the implications of an open marketplace in western Canadian wheat,durum and barley for farmers[EB/OL].2008.http://www.agric.gov.ab.ca/$Department/deptdocs.nsf/all/choice13151/$FILE/informafinalreport.pdf.
[6] DePauw R M,Hunt T.Canadian wheat pool[M]//Bonjean A P,Angus W J.The world wheat book.A history of wheat breeding.Secaucus,NJ:Lavoisier Publishing Inc.,2001:479-515.
[7] McCallum B D,DePauw R M.A review of wheat cultivars grown in the Canadian Prairies[J].Can J Plant Sci,2008(88):649-677.
[8] Dexter J E,Preston K R,Woodbeck N J.Canadian Wheat[M]//Popper L,Schafer W,Freund W.Future of flour a compendium of flour improvement,Dunne:Agrimedia,Bergen,2006:43-62.
[9] McCallum B D,Seto-Goh P,Xue A G.Physiologic specialization of Puccinia triticina in Canada in 2007[J].Can J Plant Pathol,2010(32):229-236.
[10] Fetch T G Jr,Mitchell Fetch J W,Xue A G.Races of physiologic specialization of Puccinia graminis on barley and wheat in Canada in 2006[J].Can J Plant Pathol,2011,33:54-60.
[11] Gilbert J,Tekauz A.Review:Recent developments in research on fusarium head blight of wheat in Canada[J].Can J Plant Pathol,2000(22):1-8.
[12] Harker K N,Clayton G W,Blackshaw R E,et al.Glyphosateresistant wheat persistence in western Canadian cropping systems[J].Weed Sci,2006,53:846-859.
[13] Goodwin M.Crop profile for wheat in Canada[EB/OL].[2005-10-16]http://publications.gc.ca/collections/collection_2009/agr/A118-10-16-2005E.pdf.
[14] Fowler D B.Winter wheat production in western Canada-Opportunities and obstacles[C].Proceedings of the 2011 Soils and Crops Workshop,Saskatoon:University of Saskatchewan,2011.
[15] Buller A H R.Essays on wheat[M].New York:The Macmillan Company,1919.
[16] DePauw R M,Hurd E A,Townley-Smith T F,et al.HY320 red spring wheat[J].Can J Plant Sci,1987,67:807-811.
[17] DePauw R M,Townley-Smith T F,McCaig T N,et al.HY355 white spring wheat[J].Can J Plant Sci,1989,69:1245-1250.
[18] Day S.The crops that will power biofuels(ethanol in particular)[C].Manitoba Agronomists Conference 2007 Proceedings,Winnipeg:University of Manitoba,2007.
[19] DePauw R M.Canadian prairie spring and Canadian western extra strong red wheat[M]//Slinkard A E,Knott D R.Harvest of gold:The history of field crop breeding in Canada.Saskatoon:Extension Press of University of Saskatchewan,1995:36-42.
[20] Bushuk W.The baking potential of Glenlea wheat[J].Can J Plant Sci,1980,60:737-739.
[21] DePauw R M,Thomas J B,Townley-Smith T F.Spring wheat production in the brown and dark brown soil zones of western Canada[M]//Slinkard A E,Fowler D B.Wheat production in Canada:A review,Proceedings of the Canadian wheat production symposium,Saskatchewan,SK,1986:27-45.
[22] Humphreys D G,Townley-Smith T F,Czarnecki E,et al.Kanata hard red spring wheat[J].Can J Plant Sci,2007,87:301-305.
[23] Humphreys D G,Townley-Smith T F,Czarnecki E,et al.Snowbird hard red spring wheat[J].Can J Plant Sci,2007b,87:301-305.
[24] Knott D R.Durum wheat[M]//Slinkard A E,Knott D R.Harvest of gold:The history of field crop breeding in Canada.Saskatoon:Extension Press of University of Saskatchewan,1995:67-81.
[25] Fowler D B,Gusta L V.Selection for winter hardiness in wheat.I.Identification of genotypic variability[J].Crop Sci,1979(19):769-772.
[26] Thomas J B,Schaalje G B,Roberts D W A.Prolonged freezing of dark-hardened seedlings for rating and selection of winter wheats for winter survival ability[J].Can J Plant Sci,1988(68):47-55.
[27] McCallum B D,Fetch T,Chong J.Cereal rust control in Canada[J].Aus J Agric Res,2007b,58:639-647.
[28] Peturson B.Wheat rust epidemics in western Canada in 1953,1954,and 1955[J].Can J Plant Sci,1958,38:16-28.
[29] Singh R P,Huerta-Espino J,Roelfs A P.The wheat rusts[M]//Curtis B C,Rajaram S,Macpherson Gómez H.Bread wheat:Improvement and production,plant production and protection Series No.30.Food and Agriculture Organization of the United Nations,Rome,Italy,2002:227-250.
[30] Singh R P,Hodson D P,Jin Y,et al.Current status,likelymigration and strategies to mitigate the threat to wheat production from race Ug99(TTKS)of stem rust pathogen[J].CAB Reviews:Perspectives in Agriculture,Veterinary Science,Nutrition and Natural Resources.2006(1):54.
[31] Fetch T G Jr.Virulence of stem rust race TTKS on Canadian wheat cultivars[J].Can J Plant Pathol,2007,29:441.
[32] Menzies J,Gilbert J.Diseases of wheat[M]//Bailey K L,Gossen B D,Gugel R K,et al.Diseases of field crops in Canada,3rd ed.The Canadian Phytopathological Society,Saskatoon:University Extension Press,2002:94-128.
[33] Roelfs A P.Epidemiology in North America[M]//Roelfs A P,Bushnell W R.Cereal rusts.II.New York:Academic Press,1985:403-434.
[34] Roelfs A P.Epidemiology of the cereal rusts in North America[J].Can J Plant Path,1989(11):86-90.
[35] Knott D R.The wheat rusts-Breeding for resistance.Monographs on theoretical and applied genetics 12[M].Berlin:Springer Verlag,1989:201.
[36] Singh R P,Huerta-Espino J,PfeifferW,et al.Occurrence and impact of a new leaf rust race on durum wheat in northwestern Mexico from 2001 to 2003[J].Plant Dis,2004,88:703-708.
[37] Stubbs R W,Prescott J M,Saari E E,et al.Cereal disease methodology manual[M].Mexico,DF,CIMMYT,1986:46.
[38] Kolmer J A.Physiologic specialization of Puccinia triticina in Canada in 1998[J].Plant Dis,2001,85:155-158.
[39] Samborski D J.Wheat leaf rust[M]//Roelfs A P,Bushnell W R.The cereal rusts.Diseases,distribution,epidemiology,and control,Orlando,FL:Academic Press,1985:39-59.
[40] Su H,Conner R L,Graf R J,et al.Virulence of Puccinia striiformis f.sp.tritici,cause of stripe rust on wheat,in western Canada from 1984-2002[J].Can J Plant Pathol,2003,25:312-319.
[41] Jin Y,Szabo L,Carson M.Century-old mystery of Puccinia striiformis life history solved with the identification of Berberis spp.as an alternate host[J].Phytopathology,2010,100:432-435.
[42] Conner R L,Kuzyk A D.Effectiveness of fungicides in controlling stripe rust,leaf rust,and black point in soft white spring wheat[J].Can J Plant Pathol,1988(10):321-326.
[43] McIntosh R A,Wellings C R,Park R F.Wheat rusts-An atlas of resistance genes[M].CSISO Australia in conjunction with Kluwer.The Netherlands:Academic Publishers,Dordrecht,1995:200.
[44] Goulden C H,Stevenson T M.Breeding for disease-resistance in Canada.Part I.Cereals[J].Empire J Experimental Agricul,1949,17:133-140.
[45] Green G J,Campbell A B.Wheat cultivars resistant to Puccinia graminis tritici in western Canada:Their development,performance,and economic value[J].Can J Plant Pathol,1979(1):3-11.
[46] Rowland G G,Kerber E R.Telocentric mapping hexaploid wheat of genes for leaf rust resistance and other characters derived from Aegilops squarrosa[J].Can J Genet.Cytol,1974,16:137-144.
[47] Townley-Smith T F,Czarnecki E M.AC Cora hard red spring wheat[J].Can J Plant Sci,2008,88:157-160.
[48] DePauw R M,Knox R E,Clarke J M,et al.Lovitt hard red spring wheat[J].Can J Plant Sci,2004,84:811-814.
[49] Hiebert C W,Thomas J B,Somers D J,et al.Microsatellite mapping of adult-plant leaf rust resistance gene Lr22a in wheat[J].Theor Appl Genet,2007,115:877-884.
[50] Dyck P L.The inheritance of leaf rust resistance in the wheat cultivar Pasqua[J].Can J Plant Sci,1993b,73:903-906.
[51] Hiebert C,Thomas J,McCallum B D.Locating the broad-spectrum wheat leaf rust resistance gene Lr52(LrW)to chromosome 5B by a new cytogenetic method[J].Theor Appl Genet,2005,110:1453-1457.
[52] McCallum B D,Chen X,Shorter S,et al.Stripe rust reaction of 28 Canadian wheat cultivars[J].Can J Plant Pathol,2007a,29:401-407.
[53] Kolmer J A,Dyck P L,Roelfs A P.An appraisal of stem and leaf rust resistance in North American hard red spring wheats and the probability of multiple mutations to virulence in populations of cereal rust fungi[J].Phytopathology,1991,81:237-239.
[54] Martens J W,Dyck P L.Genetics of resistance to rust in cereals from a Canadian perspective[J].Can J Plant Pathol,1989,11:78-85.
[55] Wanyera R,Kinyua M G,Jin Y,et al.The spread of stem rust caused by Puccinia graminis f.sp.tritici,with virulence on Sr31 in wheat in Eastern Africa[J].Plant Dis,2006,90:113.
[56] Fetch T G Jr,Jin Y.Potential impact of a widely virulent stem rust race in Kenya on Canadian wheat production[J].Can J Plant Pathol,2005,27:468.
猜你喜欢
南方农业学报(2022年11期)2022-03-18
湖北农业科学(2021年10期)2021-06-25
世界农药(2020年4期)2020-12-22
农业科技与信息(2020年20期)2020-12-18
农业知识(2019年7期)2019-01-06
现代园艺(2017年22期)2018-01-19
植物保护(2017年4期)2017-08-09
浙江农业学报(2017年1期)2017-05-17
中国马铃薯(2017年1期)2017-03-02
中国茶叶(2016年4期)2016-01-18
