
中国马铃薯晚疫病菌生理小种研究进展
2017-03-02 09:15孙继英汝甲荣
中国马铃薯 2017年1期
王 腾,马 爽,孙继英,汝甲荣
(黑龙江省农业科学院克山分院,黑龙江 克山 161606)
综述
中国马铃薯晚疫病菌生理小种研究进展
王 腾*,马 爽,孙继英,汝甲荣
(黑龙江省农业科学院克山分院,黑龙江 克山 161606)
由致病疫霉(Phytophthora infestans)引起的晚疫病是马铃薯生产中最具毁灭性的病害,生理小种是晚疫病菌重要的表现型之一,其组成与变异直接关系到马铃薯晚疫病的发生与流行。对马铃薯晚疫病菌生理小种的研究进展进行了综述,分析了中国晚疫病菌生理小种的多样性及生理小种毒力基因的复杂性,并指出了目前中国晚疫病菌生理小种研究存在的问题,为今后马铃薯晚疫病菌生理小种的研究提供思路。
马铃薯;致病疫霉;生理小种
马铃薯(Solanum tuberosum L.)是世界第四大粮食作物,中国马铃薯生产面积与总产均居世界首位,是世界马铃薯第一生产大国。中国已于2015年启动马铃薯主粮化战略,马铃薯将成为中国继水稻、小麦、玉米后又一主粮。预计2020年50%以上的马铃薯将作为主粮消费[1]。由致病疫霉(Phytophthora infestans)引起的晚疫病(Late blight)是对世界马铃薯生产危害最为严重的病害。由晚疫病造成的损失一般年份减产也在20%左右,严重时减产50%以上,甚至绝产。
生理小种也被称为致病型(Pathotype),是晚疫病菌重要的表现型之一,其组成与变异直接关系到马铃薯晚疫病的发生与流行[2,3]。马铃薯晚疫病菌生理小种类型的确定,是利用一套分别具有r及R1~R11单基因的12个晚疫病鉴别寄主进行活体接种鉴定,然后根据晚疫病菌在鉴别寄主上的发病情况确定病原菌的生理小种类型。中国自20世纪60年代开始对马铃薯晚疫病菌生理小种的研究,近年来,随着各地A2交配型的发现以及地区间的频繁调种,晚疫病菌生理小种复杂程度日益增大,但各地区间、年际间生理小种复杂性的比较多停留于直观感受上,缺少指数性的分析。
1 国外马铃薯晚疫病菌生理小种研究概述
马铃薯晚疫病菌的生理分化现象首次发现于1919年,随后世界各地发现了一些生理小种。Black等[4]在1953年提出了马铃薯晚疫病菌生理小种的命名方案,利用含有r、R1、R2、R3和R4基因的16个鉴别寄主进行生理小种的鉴定。随后,世界普遍使用Black系统进行生理小种的鉴定[5-11]。随着研究的深入,又陆续发现更多的R基因[12-15],并逐步建立起了利用11种单抗性基因鉴别寄主鉴定晚疫病生理小种的方法,并沿用至今。对马铃薯晚疫病菌的生理分化现象的称呼也几经变迁,自马铃薯晚疫病菌的生理分化现象发现以来,对这种现象一直存在生理小种与致病型2种称呼,经过长期深入研究,目前国际上多称之为致病型。
1984年Gurtler[16]分析了丹麦晚疫病菌的生理小种组成情况,从来自丹麦16个不同地区分离的25个菌株中鉴定出11个不同的生理小种类型,存在7个毒力基因分别为1、3、4、5、7、10和11,优势小种为1.3.4.10。1989年Tooley和Therrien[17]鉴定了秘鲁的生理小种,发现秘鲁最为常见的晚疫病菌生理小种类型为0、1和1.5,其频率分别为33%、39%和15%。1994年Andrivon[18]在1991~1993年自法国采集的116个晚疫病菌株中共发现19个生理小种类型,1991年优势小种为1.3.4.7.11、1.3.4.7.8.11和1.3.4.7.10.11,1992和1993年则分别以小种1.3.4.7和2.3.4.7.10.11为主。1998年Peters和Platt[19]从采集自加拿大的80株晚疫病菌株中鉴定出28个生理小种和11种毒力基因,且发现1994~1996年加拿大的晚疫病菌生理小种所含毒力基因的数量呈现增加趋势,病菌的致病力也表现为逐年提高,致使许多抗病品种丧失抗病性。1996年Sujkowski等[20]鉴定了1985~1991年采集自波兰的95个晚疫病菌株,共鉴定出22个生理小种类型,优势小种为1.2.3.4.7.10.11。Ghimire等[21]对1999~2000年采自尼泊尔的251个晚疫病菌株鉴定生理小种,共发现30个生理小种类型,优势小种为4.11,生理小种多样性指数Gleason指数与Shannon指数分别为5.24、2.59,生理小种毒力基因复杂性指数Ci=2.37,Cp=3.30。2002年Knapova和Gisi[22]分析了法国和瑞典的晚疫病菌生理小种类型,以小种1.3.4.7.10.11、1.3.4.6.7.10.11、1.3.4.7.8.10.11和1.3.4.6.7.8.10.11为主,上述4个生理小种的频率之和达到91%。2002年Cohen[23]对1983~2000年以色列晚疫病菌生理小种组成情况进行了分析,在1983~1991年采集的80个晚疫病菌株中鉴定出5个生理小种类型,其中,优势小种1.3.4.7.8.10的频率为76%;在1993~1998年采集的173个晚疫病菌株中鉴定出19个生理小种,优势小种为1.3.4.7.8.10.11,其频率为63%;在1999~2000年采集的71个晚疫病菌株中鉴定出42个生理小种类型,其中就有34个为新出现的小种类型。2009年Runno-Paurson等[24]对2002~2003年采集自爱沙尼亚的101个晚疫病菌株进行生理小种鉴定,优势小种为1.3.4.7.8.10.11和1.3.4.7.10.11。Runno-Paurson等[25]从2004~2005年采集自爱沙尼亚的196个晚疫病菌株中鉴定出38个生理小种类型,优势小种为1.3.4.7.10.11,并发现3株“超级毒力小种”。Runno-Paurson等[26]从2001~2007年采集自爱沙尼亚东部的180个晚疫病菌株中鉴定出70个生理小种类型,优势小种为1.2.3.4.7.8.10.11和1.2.3.4.6.7.10.11,但频率较低(18%),于2005年发现3株“超级毒力小种”,Shannon多样性指数最高的年份为2003年(1.0)。Runno-Paurson等[27]对2011~2012年采集自立陶宛的70株晚疫病菌株进行了致病型鉴定,在2011年的40个菌株中鉴定出29种致病型,优势小种为1.2.3.4.5.6.7.8.9.10.11,并发现4株“超级毒力小种”,在2012年的30个菌株中鉴定出15种致病型,优势小种为1.2.3.4.6.7.10.11,并发现3株“超级毒力小种”。Aav等[28]在2010~2012年采自拉脱维亚的156个晚疫病菌株中鉴定出69个生理小种类型,优势小种为1.2.3.4.7.10.11与1.2.3.4.6.7.10.11,Shannon多样性指数为0.73。Michalska等[29]对2005~2007年从波兰马铃薯与番茄上采集的176个晚疫病菌株进行生理小种鉴定,共鉴定出44个生理小种类型,其中12个生理小种为马铃薯与番茄共有的,从马铃薯上鉴定出14个生理小种类型,优势小种为1.2.3.4.5.6.7.8.10.11与 1.2.3.4.5.6.7.10.11,并发现7株“超级毒力小种”,生理小种毒力基因的复杂性表明2006年复杂程度最高。
上述研究结果表明,自20世纪80年代以来,世界各国的生理小种组成均趋于复杂化,生理小种类型由单基因小种向多基因小种发展,优势小种的毒力基因组成日趋复杂。
2 中国马铃薯晚疫病菌生理小种研究进展
中国的晚疫病生理小种研究开始于20世纪60年代,张明厚[30]综合国外对马铃薯晚疫病菌生理小种的研究进展,对生理小种的分类与命名、世界各地生理小种的分布、生理小种的形成与变异以及生理小种的鉴定方法做出介绍。1981年黄河等[31]首次报道了中国马铃薯晚疫病菌生理小种的组成,但受限于当时试验条件和研究条件的限制,最初所采用的鉴别寄主较少,较单一,仅能鉴别出部分毒力基因。其后随着研究的深入,马铃薯晚疫病菌生理小种的研究才逐步成熟。根据中国晚疫病菌生理小种的研究历史,可将中国研究马铃薯晚疫病菌生理小种的历史分为3个阶段。
第一阶段,研究的初始阶段。中国马铃薯晚疫病菌生理小种的研究始于20世纪60年代,黄河等[31]对1962~1967年从张家口、黑龙江、甘肃、山西、内蒙、青海和北京等地收集的晚疫病菌进行了生理小种鉴定,鉴定出7个生理小种类型,并发现生理小种出现由简单变复杂又趋于简单的情况。随后,李克来[32]对1969~1981年采集自呼和浩特的马铃薯晚疫病菌株进行了生理小种鉴定,共鉴定出6个生理小种类型,小种1为优势小种。之后,又再次鉴定了1969~1985年采集的晚疫病菌株[33],发现在晚疫病严重发生的年份生理小种出现的种类增多,多毒力基因小种容易出现,在晚疫病少发病的干旱年份生理小种出现的种类较少。刘晓鹏等[34]采用了不含R10与R11的10个单基因鉴别寄主及多基因鉴别寄主对湖北恩施地区进行了生理小种鉴定,但其生理小种的命名是不对的。姚裕琪和梁德霖[35]利用7份含单基因及8份含多基因的鉴别寄主从采集自内蒙古的晚疫病菌中共鉴定出1、3、4、1.3、2.3和3.4共6个生理小种,但鉴别寄主所含基因不明确。杨艳丽等[36]对采自云南的41株晚疫病菌株进行生理小种鉴定,共鉴定出0、3、4及3.4共4个生理小种。Ryu等[37]在2000年采自云南的菌株中鉴定出0、3、4和3.4共4个生理小种类型。陈茜[38]从2006~2007年采集自四川省的131个晚疫病菌株中鉴定出8个生理小种类型,优势小种为3.4。刘波微等[39]从2003~2007年采集自四川的241个晚疫病菌株中共鉴定出11个生理小种类型,优势小种为3.4。以上研究在生理小种鉴定上存在2个不足:(1)采用的鉴别寄主不完全,鉴别寄主不论是含单基因还是复合基因,其结果均只能鉴别出毒力基因R1、R2、R3与R4;(2)部分学者的研究对生理小种的命名存在错误。
第二阶段,研究的发展阶段。随着国际合作的加深及对晚疫病菌生理小种研究的深入,中国学者逐步从国际马铃薯中心引进了能鉴别所有11个毒力基因的全部鉴别寄主。但在研究中应用的鉴别寄主仍不完全(表1),尤其缺少能够鉴定毒力基因类型的单基因鉴别寄主,因此不能准确说明生理小种所具有的毒力基因。这种生理小种鉴定结果缺乏严格性,应用中有其局限性。
第三阶段,研究的成熟阶段。随着研究的继续深入及科研条件的改善,中国马铃薯晚疫病菌生理小种的研究逐步成熟,中国学者对马铃薯晚疫病菌生理小种进行了大量研究。杨素祥[47]在2002~2005年采自云南的155株晚疫病菌株中鉴定出55个生理小种类型,优势小种为1.2.3.4.5.6.7.8.9.10,频率为32.89%,且发现生理小种的分布没有明显区域性;杨艳丽等[48]对2003~2005年自云南采集的117个晚疫病菌株进行生理小种类型的鉴定,共鉴定出26个生理小种类型,占主导的小种为3.4.6.8.10.11,其频率为28.69%。郭军等[49]从1997~1999年和2002~2003年自内蒙古采集的38个晚疫病菌株中鉴定出18个生理小种类型,小种3a.3b.4.7.10频率最高,占所测菌株的21.05%,小种1.2.3a.3b.4.5.6.7.8.9.10.11占13.16%,小种3a.3b.4.6.7.10占10.53%。赵志坚等[50]对2000~2003年自云南省采集的136个晚疫病菌株鉴定生理小种类型,共鉴定出28个生理小种类型,主要小种类型为1.3.4.7.9.10.11、1.2.3.4.6.7.9.10.11、1.3.4.6.7.9.10.11以及3.4.6.9.10.11,且发现1株含有已知11个毒力基因的“超级毒力小种”。2008年叶广继[51],叶广继等[52]以及孙海宏和王舰[53]分别对2006~2007年采集自青海的晚疫病菌株进行生理小种鉴定,其结果均显示青海省马铃薯晚疫病菌优势小种为3.4.10。金光辉等[54]对2008年采自黑龙江的83个马铃薯晚疫病菌株进行了生理小种鉴定,共鉴定出18个生理小种类型,优势小种为3.4.7.11,小种3.4.5.6.7.8.9.10.11含有的毒力基因数目最多。李洪浩等[55]从2008~2009年采自四川的103个晚疫病菌株中鉴定出42个生理小种类型,优势小种为1.2.3.4.5.6.7.8.10.11,并发现5株“超级毒力小种”。桂春爽[56]从2008~2009年采集自河北、黑龙江、辽宁和吉林的150个晚疫病菌株中鉴定出20个生理小种类型,其中河北采集的59个菌株中鉴定出11个生理小种类型,76个采自黑龙江的菌株中鉴定出15个生理小种类型,两省的优势小种均为“超级毒力小种”。韩彦卿等[57]在2006~2008年采自河北、黑龙江、福建、四川、云南和内蒙古6个省份的57个晚疫病菌株中,共鉴定出30个生理小种类型,并发现3株超级毒力小种。刘狄[58]对2009年采自湖北的55个晚疫病菌株进行生理小种鉴定,共鉴定出29个生理小种类型,并发现5株“超级毒力小种”。成兰芳等[59]以及成兰芳和王生荣[60]从2009年甘肃省采集的63个晚疫病菌株中鉴定出16个生理小种类型,占主导的小种4.6.7.11的频率达到28.57%。王鹤[61]从2009年自黑龙江、吉林、云南和内蒙古四省(区)采集的118个晚疫病菌株中鉴定出42个生理小种类型,在黑龙江34个菌株中鉴定出25个生理小种类型,在被测49个吉林菌株中鉴定出19个生理小种类型,在云南采集的24个菌株中鉴定出10个生理小种类型,在内蒙古采集的11个菌株中鉴定出6个生理小种类型,另外,还发现了35株“超级毒力小种”,且在各省均有发现。王晨[62]从2008~2011年采集自黑龙江的41个晚疫病菌株中鉴定出15个生理小种类型,优势小种为1.2.3.4.5.6.7.9.10.11,并发现5株“超级毒力小种”。郭梅[63]对2010年采自黑龙江的95个晚疫病菌株进行生理小种鉴定,共鉴定出24个生理小种类型,优势小种为1.3.4.7.8.10.11。Wang等[64]对2004~2008年采集自黑龙江的72个晚疫病菌株进行生理小种鉴定,共鉴定出39个生理小种类型,优势小种为1.3.4.7.10.11。高云飞等[65]从2010年自哈尔滨市采集的62个晚疫病菌株中鉴定出14个生理小种类型,优势小种1.3.4.7.8.10.11出现频率为33.87%。马云芳等[66]在2009年自宁夏从1个种质资源圃采集的43个晚疫病菌株中鉴定出2个小种类型1.2.3.4.5.6.7.8.9.10.11和3.4.10,频率分别为95.3%、4.7%,但因取样范围过小,代表性不好。李洪浩等[67]从2008~2011年自四川采集的192个晚疫病菌株中共鉴定出55个生理小种类型,优势小种为“超级毒力小种”,仅发现1株单毒力基因小
种。王立等[68]在自甘肃省采集的75个菌株中鉴定出45个生理小种类型,其中小种l.2.3.4.5.6.7.8.9.10.11为优势小种,频率为22.67%。李继平[69]在2007~2012年采自甘肃的77个晚疫病菌株中共鉴定出45个生理小种类型,优势小种为“超级毒力小种”。韩淼等[70]对2007~2008年采自甘肃和内蒙古地区的晚疫病菌株鉴定生理小种,在来自甘肃的65个菌株中鉴定出28个小种类型,有3株“超级毒力小种”;在来自内蒙古的26个菌株中,共鉴定出24个小种类型,有2株“超级毒力小种”。覃雁瑜[71]对2012~2013年采集自云南的晚疫病菌株进行生理小种鉴定,从2012年采集的292个菌株中鉴定出99个生理小种类型,从2013年采集的205个菌株中鉴定出27个生理小种类型,2年的优势小种均为“超级毒力小种”。陈晓梅[72]对2010~2011年采集自甘肃、宁夏、云南、福建、霞浦、贵州、南宁等地的菌株中每地选取20个菌株进行生理小种鉴定,各地均发现17个以上的生理小种类型,发现6株“超级毒力小种”,并进行了生理小种多样性及生理小种毒力基因复杂性分析。王腾[73]对2010~2013年采集自黑龙江的208个晚疫病菌株进行生理小种鉴定,共鉴定出75个生理小种类型,在2012与2013年分别发现2株与1株“超级毒力小种”,进行了生理小种多样性及生理小种毒力基因复杂性分析,发现2012年生理小种复杂性及生理小种毒力基因复杂性均最高。杨胜先等[74]对2013~2014年采集自贵州的260个晚疫病菌株进行生理小种鉴定,其中2013年鉴定出15个生理小种类型,2014年鉴定出16个生理小种类型,2年优势小种均为2.5.6.8.9.11,未发现“超级毒力小种”。
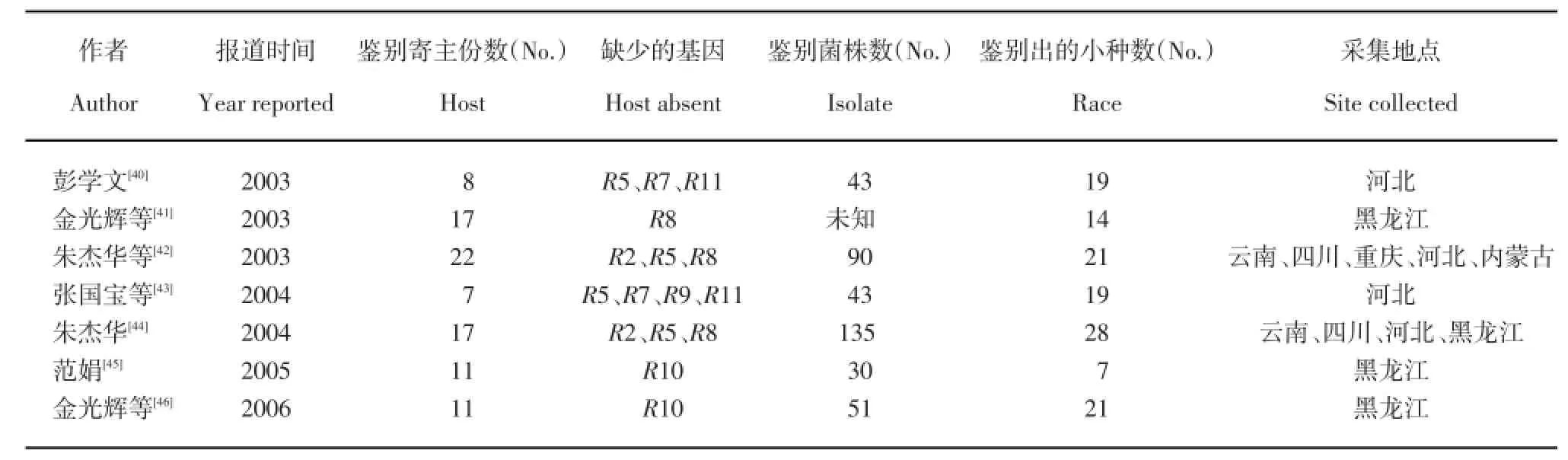
表1 中国马铃薯晚疫病菌生理小种的鉴定Table 1 Race of Phytophthora infestans identified on potato in China
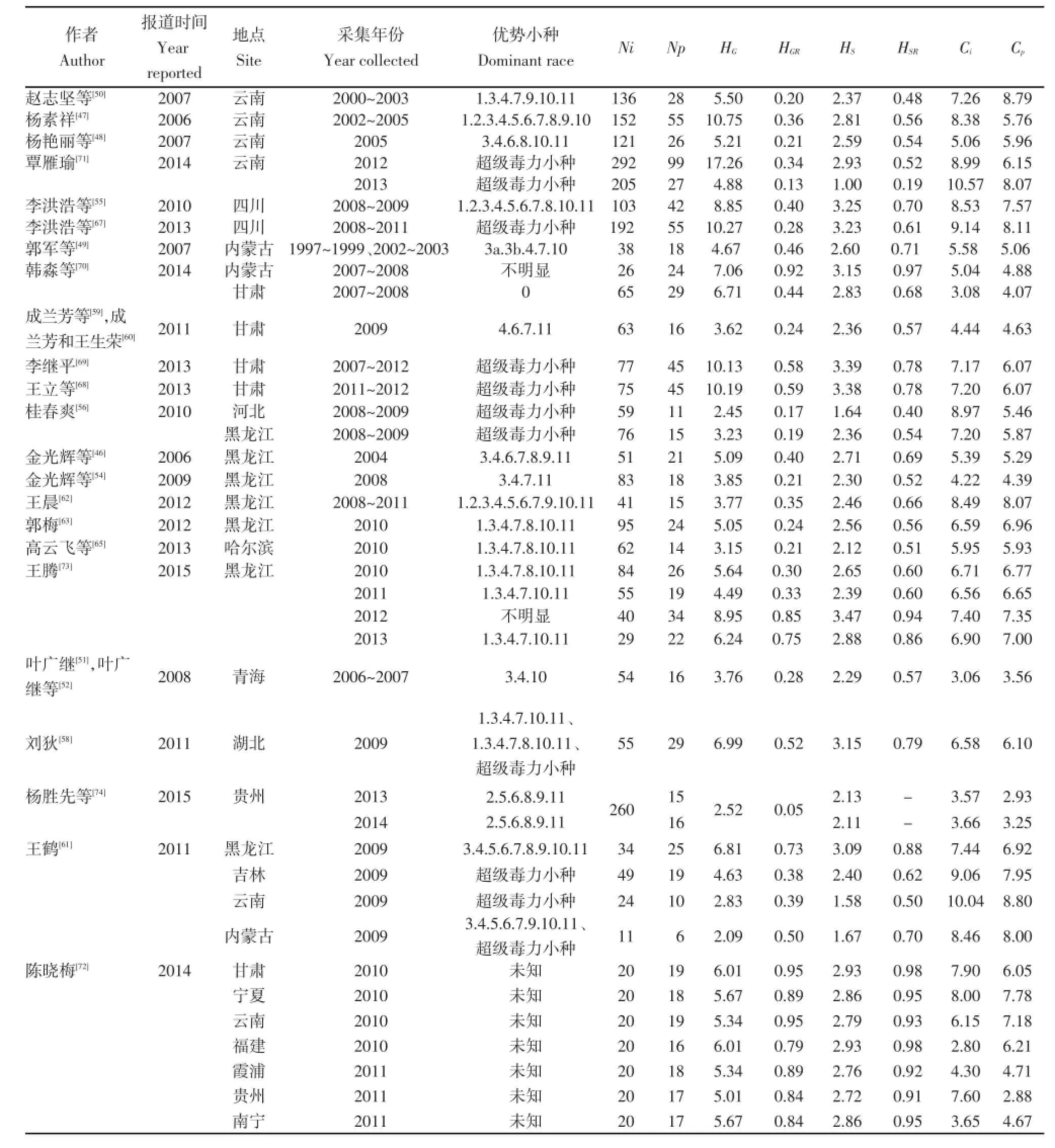
表2 中国马铃薯晚疫病菌生理小种多样性及生理小种毒力基因复杂性Table 2 Race diversity and virulence complexity of Phytophthora infestans on potato in China
由这些研究看,近年来,“超级毒力小种”在各省的出现越来越普遍,部分省份甚至“超级毒力小种”已成为优势小种。从马铃薯晚疫病生理小种的多样性(HG、HGR、HS、HSR)看,各省并没有明显的普遍规律;从生理小种毒力基因的复杂性(Ci、Cp)看,各省总体上均表现出升高的趋势(表2)。这说明各省的晚疫病菌每个菌株及每个生理小种所含毒力基因数目较以前更多,这必将导致马铃薯品种对晚疫病抗性的降低。马铃薯晚疫病菌生理小种的这些变化使马铃薯晚疫病的防治难度变大,对马铃薯抗晚疫病育种及防治晚疫病提出了更加严峻的挑战。
3 中国生理小种研究存在的问题及展望
中国对马铃薯晚疫病菌生理小种的研究已进行了大量工作,研究方法也日渐成熟,但仍存在一些问题需要改进。
研究连续性不强。在晚疫病生理小种的研究中,部分省份缺乏连续性,如云南省2005~2010年中断4年,内蒙古2003~2007年中断3年,2008年至今未见报道,四川2011年至今未见报道。另外,部分省份病样采集地点在年际间存在差异,有的年份采集地点过少。而这必将造成无法明确生理小种随时间变化的规律,以及无法确定生理小种出现变化的原因。
对生理小种多样性及毒力基因复杂性缺少客观的指数性分析。至今中国对晚疫病菌生理小种的指数性分析仅有陈晓梅[72]及王腾[73]报道过,但均未应用相对指数HGR及HSR,因此对生理小种多样性的比较存在局限性。
虽然目前中国对马铃薯晚疫病菌生理小种的研究存在一定的问题和局限性,但随着研究人员对晚疫病菌生理小种的认识及国际合作的加强,相信中国马铃薯晚疫病菌生理小种的研究会更加成熟。
[1] 付承堃.中国将启动马铃薯主粮化战略土豆或成第四主粮[N].国际在线,2015-01-06.
[2] Koh Y J,Goodwin S B,Dyer A T,et al.Migrations and displacements of Phytophthora infestans populations in East Asian countries[J].Phytopathology,1994,84(9):922-927.
[3] 朱杰华,杨志辉,张凤国,等.马铃薯晚疫病菌群体遗传结构研究进展[J].中国农业科学,2007,40(9):1936-1942.
[4] Black W,Mastenbroek C,Mills W R,et al.A proposal for an internationalnomenclature of races of Phytophthora infestans and of genes controlling immunity in Solanum demissum derivatives[J]. Euphytica,1953,2(3):173-179.
[5] Mastenbroek C,Bruin T D.Het voorkomen van physio 4 van Phytophthora infestans in Netherlands[J].European Journal of Plant Pathology,1955,61(1):88-92.
[6] Howatt J L,Grainger P N.Some new findings concerning Phytophthora infestans(Mont.)De By[J].American Journal of Potato Research,1955,32(5):180-188.
[7] Graham K M.Distribution of physiological races of Phytophthora infestans(Mont.)de Bary in Canada[J].American Journal of Potato Research,1955,32(8):277-282.
[8] Pristou R,Gallegly M E.Differential reaction of potato hosts to foreign and domestic potato physiologic races of Phytophthora infestans[J].American Potato Journal,1956,33(10):287-295.
[9] Gallegly M E,Eichenmuller J J.The spontaneous appearance ofthe potato race 4 character in cultures of Phytophthora infestans[J]. American Potato Journal,1959,36(2):45-51.
[10] Graham K M,Niederhauser J S,Romero S.Observations on races of Phytophthora infestans in Mexico during 1956-1957[J]. American Journalof Potato Research,1959,36(6):196-203.
[11] Mastenbroek C.Some major points from 22 years of experience in breeding potatoes for resistance to late blight(Phytophthora infestans)[J].American Potato Journal,1966,43(8):261-277.
[12] Schick R,Schick E,Haussdorfer M.Ein Beitrag zur physiologischen spezialisierung von Phytophthora infestans[J]. Phytopath Z,1958,31:225-236.
[13]Eide C J,Bonde R,Gallegly M E,et al.Report of the late blight investigations committee[J].American Potato Journal,1960,37 (11):399-400.
[14] Mckee R K.Identification of R-genes in Solanum stoloniferum[J]. Euphytica,1962,11(1):42-46.
[15] Malcolmson J F,Black W.New R genes in Solanum demissum Lindl.and their complementary races of Phytophthora infestans (Mont.)de Bary[J].Euphytica,1966,15(2):199-203.
[16] Gurtler H.Physiological races of Phytophthora infestans in Denmark and low temperature storage of isolates[J].Potato Research,1984,27(1):25-31.
[17] Tooley P M,Therrien D L.Mating type,race composition,nuclear DNA content,and isozyme analysis of Peruvian isolates of Phytophthora infestans[J].Phytopathology,1989,79(4):478-481.
[18] Andrivon D.Races of Phytophthora infestans in France, 1991-1993[J].Potato Research,1994,37(3):279-286.
[19] Peters R D,Platt H W.Changes in race structure of Canadian populations of Phytophthora infestans based on specific virulence to selected potato c1ones[J].Potato Research,1998,41(4): 366-370.
[20] Sujkowski L S,Goodwin S B,Fry W E.Changes in specific virulence in Polish populations of Phytophthora infestans: 1985-1991[J].European Journal of Plant Pathology,1996,102 (6):555-561.
[21] Ghimire S R,Hyde K D,Hodgkiss I J,et al.Race diversity and virulence complexity of Phytophthora infestans in Nepal[J].Potato Research,2001,44(3):253-263.
[22] Knapova G,Gisi U.Phenotypic and genotypic structure of Phytophthora infestans populations on potato and tomato in France and Switzerland[J].Plant Pathology,2002,51(5):641-653.
[23]Cohen Y.Populations of Phytophthora infestans in lsrael underwent three major genetic changes during 1983 to 2000[J].Phytopathology, 2002,92(3):300-307.
[24] Runno-Paurson E,Fry W E,Myers K L,et al.Characterisation of Phytophthora infestans isolates collected from potato in Estonia during 2002-2003[J].European Journalof Plant Pathology,2009, 124(4):565-575.
[25]Runno-Paurson E,Remmel T,Ojarand A,et al.The structure of Phytophthora infestans populations from organic and conventional crops[J].Euopeanr Journal of Plant Pathology,2010,128(3): 373-383.
[26]Runno-Paurson E,Hannukkala A,Trdan S,et al.The structure of mating type,virulence,metalaxyl resistance of Phytophthora infestans in a long-term phenotypic study in distinct location in Eastern Estonia[J].Journal of Plant Diseases and Protection, 2012,119(2):45-52.
[27] Runno-Paurson E,Ronis A,Hansen1 M,et al.Lithuanian populations of Phytophthora infestans revealed a high phenotypic diversity[J].Journal of Plant Diseases and Protection,2015,122 (2):57-65.
[28] Aav A,Skrabule I,Bimšteine G,etal.The structure ofmating type, metalaxyl resistance and virulence of Phytophthora infestans isolates collected from Latvia[J].Zemdirbyste-agriculture,2015, 102(3):335-342.
[29] Michalska A M,Sobkowiak S,Flis B,et al.Virulence and aggressiveness of Phytophthora infestans isolates collected inPoland from potato and tomato plants identified no strong specificity[J].European Journal of Plant Pathology,2015,144(2): 325-336.
[30] 张明厚.马铃薯晚疫病菌(Phytophthora infestans(Mont.)de Bary)生理分化现象的研究概况[J].东北农学院学报,1964(1):33-46.
[31] 黄河,程汉清,徐天宇,等.我国北部马铃薯晚疫病菌生理小种的发生和变化[J].植物病理学报,1981,11(1):47-51.
[32] 李克来.Phytophthora infestans在番茄上的特异性及马铃薯致病性的关系 [J].云南师范大学学报:自然科学版,1987(2): 44-51.
[33]李克来.呼和浩特马铃薯晚疫病病菌生理小种组成及块茎生理小种毒性与植株感染关系[J].内蒙古大学学报:自然科学版, 1988(1):182-187.
[34]刘晓鹏,谢从华,宋伯符.湖北恩施地区马铃薯晚疫病病菌生理小种的组成及分布[J].马铃薯杂志,1995,9(2):81-83.
[35] 姚裕琪,梁德霖.内蒙古马铃薯晚疫病病菌生理小种的组成及分布[J].内蒙古农业科技,1996(1):17-18.
[36] 杨艳丽,罗文富,国立耘.云南马铃薯晚疫病菌生理小种的研究[J].植物保护,2001,27(4):3-5.
[37] Ryu K Y,Luo W F,Yang Y L,et al.Mating type,fungicide sensitivity and physiological race of Phytophthora infestans collected from Yunnan Province[J].Acta Phytopathologica Sinica, 2003,33(2):126-131.
[38] 陈茜.四川省马铃薯晚疫病菌生物学特性与遗传多样性研究[D].雅安:四川农业大学,2008.
[39] 刘波微,彭华贤,席亚东,等.四川马铃薯晚疫病生理小种鉴定及品种抗病评价[J].西南农业学报,2010,23(3):747-751.
[40] 彭学文.河北省马铃薯病害调查及主要真菌病害研究[D].保定:河北农业大学,2003.
[41] 金光辉,文景芝,董传民,等.黑龙江省马铃薯晚疫病生理小种的类型与分布状况研究[J].中国马铃薯,2003,17(4):213-215.
[42] 朱杰华,杨志辉,邵铁梅,等.中国部分地区马铃薯晚疫病菌生理小种的组成及分布[J].中国农业科学,2003,36(2):169-172.
[43]张国宝,朱杰华,彭学文.河北省部分地区马铃薯晚疫病菌生理小种鉴定[J].河北农业大学学报,2004,27(1):77-79.
[44] 朱杰华.中国马铃薯晚疫病菌群体遗传结构研究[D].保定:河北农业大学,2004.
[45] 范娟.马铃薯抗晚疫病资源筛选的研究[D].哈尔滨:东北农业大学,2005.
[46] 金光辉,袁善奎,吕文河,等.黑龙江省马铃薯晚疫病菌生理小种类型及交配型研究[J].东北农业大学学报,2006,37(5): 610-614.
[47] 杨素祥.云南马铃薯晚疫病菌群体的遗传多样性研究[D].昆明:云南师范大学,2006.
[48] 杨艳丽,胡先奇,鲁绍凤,等.云南省马铃薯晚疫病菌生理小种的组成与分布[J].华中农业大学学报,2007,26(3):297-301.
[49] 郭军,屈冬玉,巩秀峰,等.内蒙古马铃薯晚疫病菌交配型和生理小种研究[J].西北农林科技大学学报:自然科学版,2007,35 (11):120-124.
[50] 赵志坚,李灿辉,曹继芬,等.云南省马铃薯致病疫霉毒性基因组成及毒力结构研究[J].中国农业科学,2007,40(3):505-511.
[51] 叶广继.青海马铃薯晚疫病菌群体遗传多样性研究[D].西宁:青海大学,2008.
[52] 叶广继,孙海宏,周云,等.青海海东地区马铃薯晚疫病菌生理小种的组成及分布[J].植物病理学报,2008,38(5):107-110.
[53] 孙海宏,王舰.青海致病疫霉生理小种组成及分布研究[J].现代农业科技,2008(11):112-113.
[54] 金光辉,吕文河,白雅梅,等.黑龙江省马铃薯晚疫病菌生理小种的鉴定[J].东北农业大学学报,2009,40(10):19-23.
[55] 李洪浩,张敏,雷高,等.四川省马铃薯晚疫病菌生理小种组成及分布[C]//中国植物保护学会.公共植保与绿色防控,2010: 77-78.
[56]桂春爽.北方四省致病疫霉致病型及拮抗菌复合发酵抑菌作用的研究[D].保定:河北大学,2010.
[57] 韩彦卿,秦宇轩,朱杰华,等.2006—2008年中国部分地区马铃薯晚疫病菌生理小种的分布[J].中国农业科学,2010,43(17): 3684-3690.
[58] 刘狄.湖北西南地区马铃薯晚疫病生理小种组成分布与抗药性分析[D].武汉:华中农业大学,2011.
[59] 成兰芳,张文解,李惠霞,等.甘肃省马铃薯晚疫病菌生理小种鉴定[J].甘肃农业大学学报,2011,46(3):65-69.
[60] 成兰芳,王生荣.甘肃马铃薯晚疫病菌毒力类型及分布初探[J].西北农业学报,2011,20(11):183-186.
[61]王鹤.2009年中国4省马铃薯晚疫病菌群体特性分析及其交配型分子标记的开发[D].保定:河北农业大学,2011.
[62]王晨.马铃薯晚疫病菌的表现型和SSR基因型分析[D].哈尔滨:东北农业大学,2012.
[63] 郭梅.黑龙江省马铃薯晚疫病菌表型特性研究[D].北京:中国农业科学院,2012.
[64] Wang X,Guo M,Min F,et al.Virulence complexity and highlevels of fungicide resistance suggest population change of Phytophthora infestans in the Heilongjiang Province of China[J]. Potato Research,2012,55(3):217-224.
[65] 高云飞,郭梅,王晓丹,等.哈尔滨市马铃薯晚疫病菌生理小种的类型[J].中国马铃薯,2013,27(3):168-171.
[66] 马云芳,孙洁平,马丽杰,等.一个马铃薯种质资源圃致病疫霉群体的分析[J].菌物学报,2013,32(5):802-811.
[67] 李洪浩,彭化贤,席亚东,等.四川马铃薯晚疫病菌交配型、生理小种、甲霜灵敏感性及mt DNA单倍型组成分析[J].中国农业科学,2013,46(4):728-736.
[68]王立,惠娜娜,李建军,等.甘肃省马铃薯主产区晚疫病菌生理小种组成与分布[J].中国蔬菜,2013(22):70-74.
[69] 李继平.甘肃马铃薯晚疫病菌群体结构及病害治理技术研究[D].兰州:甘肃农业大学,2013.
[70] 韩淼,汪晓雯,黄琛,等.甘肃和内蒙古地区马铃薯晚疫病菌的致病型[J].植物保护,2014,40(2):146-150.
[71] 覃雁瑜.品种多样性对马铃薯晚疫病菌群体遗传结构的影响[D].福州:福建农林大学,2014.
[72]陈晓梅.马铃薯致病疫霉生理小种多样性及其空间分布[D].福州:福建农林大学,2014.
[73]王腾.黑龙江省马铃薯晚疫病菌群体结构研究及块茎抗病性鉴定[D].大庆:黑龙江八一农垦大学,2015.
[74] 杨胜先,张绍荣,龙国,等.贵州省马铃薯晚疫病菌生理小种的组成与分布[J].南方农业学报,2015,46(4):597-601.
[75] Groth J V,Roelfe A P.The concept and measurement of phenotypic diversity in Puccinia graminiis on wheat[J]. Phytopathology,1987,77(10):1395-1399.
[76] Andrivon D.Race structure and dynamics in populations of Phytophthora infestans[J].Canadian Journal of Botany,1994,72 (11):1681-1687.
Research Progress in PhysiologicalRace of Phytophthora infestans in China
WANG Teng*,MA Shuang,SUN Jiying,RU Jiarong
(Keshan Branch,Heilongjiang Academy ofAgriculturalSciences,Keshan,Heilongjiang 161606,China)
ract:Late blight caused by Phytophthora infestans is the most devastating disease in potato production. Physiological race of P.infestans is one of the important phenotypes,and its composition and variation are directly related to the incidence and prevalence of potato late blight.Research progress in physiologicalrace of P.infestans was reviewed.The diversity of physiological race of P.infestans and complexity of the physiological race virulence genes were analyzed,and problems relative to P.infestans physiological races were pointed out,which might help future researches on P.infestans physiologicalraces in China.
ords:potato;Phytophthora infestans;physiologicalrace
S532
A
1672-3635(2017)01-0045-09
2016-06-30
齐齐哈尔市攻关项目(NYGG-201411)。
王腾(1989-),男,研究实习员,硕士,从事马铃薯抗病育种研究。
王腾,E-mail:wteng1129@126.com。
猜你喜欢
作物学报(2021年2期)2021-12-24
检验医学(2021年11期)2021-11-29
今日农业(2021年14期)2021-10-14
今日农业(2021年7期)2021-07-28
今日农业(2020年15期)2020-09-25
山西农业大学学报(自然科学版)(2020年1期)2020-03-04
农药科学与管理(2019年9期)2019-11-23
农药科学与管理(2019年6期)2019-11-23
中国抗生素杂志(2019年6期)2019-07-06
今日农业(2019年14期)2019-01-04
