
安徽省稻瘟病菌生理小种鉴定分析
2022-03-18 05:55:26周坤能夏加发王元垒张彩娟马廷臣张爱芳李泽福
南方农业学报 2022年11期
周坤能,夏加发,王元垒,云 鹏,张彩娟,马廷臣,张爱芳,李泽福*
(1安徽省农业科学院水稻研究所/安徽省水稻遗传育种重点实验室,安徽 合肥 230001;2安徽省农业科学院植物保护与农产品质量安全研究所,安徽 合肥 230001)
0 引言
【研究意义】稻瘟病是水稻主要病害之一,每年均有大面积发生,严重威胁水稻生产(王才林等,2008;阮宏椿等,2017)。选育抗病品种是防治稻瘟病最经济有效的措施,抗病基因和材料是稻瘟病抗性育种的前提和基础(张礼霞等,2017;张柱坚等,2018;徐志健等,2020)。然而,稻瘟病属真菌性病害,生理小种易发生变异,且不同区域的稻瘟病菌结构差异显著,导致品种间的抗性差异较大(曹妮等,2019)。因此,明确稻瘟病菌生理小种类型对抗病基因挖掘和品种选育具有重要意义。【前人研究进展】利用7个水稻鉴别品种对江苏省2001—2010年的稻瘟病菌进行分析研究得出江苏省稻瘟病菌优势种群为ZG和ZB群(刘永锋等,2010);相同的方法研究表明广西稻瘟病菌优势种群为ZB群,优势生理小种为ZB9和ZB13(陈小林等,2017)。湖北省稻瘟病重发区优势种群为ZA、ZC和ZB群,优势生理小种为ZA1和ZB1;但湖北随州地区稻瘟病菌优势种群为ZB群,优势生理小种为ZB15(杨小林等,2016;吴尧等,2021)。黑龙江省稻瘟病菌优势种群为ZA和ZD群,优势小种为ZA49、ZD1和ZA1(高红秀等,2017)。广东省稻瘟病菌优势种群为ZB和ZC群,优势生理小种为ZB13、ZB15和ZC15(钟宝玉等,2018)。福建省稻瘟病菌优势种群为ZA群,ZB、ZC和ZD群亦存在较高的分布频率(邓云等,2020;2021)。山东省稻瘟病菌优势种群为ZA和ZC群,其次为ZB和ZE群,优势生理小种为ZE3和ZG1(房文文等,2020)。辽宁省稻瘟病菌优势种群为ZA和ZB群,优势生理小种为ZA1和ZB1(刘伟等,2020)。湖南省桃江病圃稻瘟病菌优势种群为ZB群,优势生理小种为ZB15和ZB1(李杰等,2021)。水稻抗稻瘟病品种在种植几年后出现抗性减弱或丧失现象,可能与稻瘟病菌生理小种的变异有关(肖丹凤等,2013)。因此,明确不同地区稻瘟病菌生理小种结构和致病力,解析抗性基因与生理小种之间的关系,有助于不同稻瘟病抗性基因的功能研究并应用于育种利用(Rathour et al.,2016;Deng et al.,2017;潘存红等,2021;Liu et al.,2021)。沈乐融等(2019)利用江苏省稻瘟病菌生理小种与10个稻瘟病抗性基因进行致病力分析,发现供试菌株对Pizt、Piz、Pita、Pita2和Pii表现为弱毒力;黑龙江省稻瘟病致病性分化研究表明,含有Pi9和Piz5的单基因系抗性较好(马军韬等,2020)。安徽省稻瘟病菌生理小种分离鉴定研究工作相对缓慢,陈莉等(2007)对2002年安徽省稻区稻瘟病菌进行生理小种鉴定,结果显示ZA~ZH群小种在安徽均有分布,优势种群为ZB群,优势生理小种为ZB13和ZB15。【本研究切入点】安徽省地理位置复杂,地势由平原、丘陵、山地构成,早晚稻、籼粳糯等类型品种均有种植,依据耕作制度、耕作方式和生产条件等将安徽省水稻种植区划分为5个稻作区,包括沿淮淮北单季稻作区,江淮丘陵单、双季稻过渡区,沿江双、单季稻作区,大别山地单、双季稻作区和皖南山地单、双季稻作区(万建民等,2018),各稻作区的稻瘟病菌生理小种结构差异较大,且各稻作区的稻瘟病菌优势种群和生理小种尚不明确。【拟解决的关键问题】利用7个水稻鉴别品种,采用苗期人工喷雾接种法对2019—2020年收集的安徽省不同稻作区稻瘟病菌进行鉴定,分析安徽省稻瘟病菌优势种群和生理小种类型以及不同稻作区稻瘟病菌结构分布,并分析优势生理小种的致病力,筛选抗稻瘟病优异种质资源,为稻瘟病抗性基因挖掘、品种选育和推广布局提供科学依据。
1 材料与方法
1.1 病穗样本采集及试验鉴别品种
2019—2020年,收集安徽省5个稻作区21个市(县、区)主栽籼稻和粳稻品种的穗颈瘟病穗样本,并记录品种名称、采集地等信息,4 ℃保存备用。7个水稻稻瘟病菌鉴别品种分别为特特普、珍龙13、四丰43、东农363、关东51、合江18和丽江新团黑谷。
1.2 稻瘟病菌分离和培养
对采集的稻瘟病病穗利用ddH2O轻微冲洗,放置于含有湿润滤纸的灭菌培养皿中保湿培养24~48 h,显微镜下观察孢子萌发情况,待长出足够孢子后,将孢子依次振落到2~3个马铃薯固体培养基上,28 ℃黑暗培养3 d,挑选单菌落于灭菌大麦中培养8~10 d,待菌丝长出后,用ddH2O洗脱菌落(周坤能等,2022)。
1.3 人工接种鉴定
制备浓度为1×105个/mL的稻瘟病菌菌液,对生长至3叶期的7个水稻鉴别品种进行喷雾接种鉴定,遮光保湿24 h后,每1 h喷水2 min保湿,3次重复。7~10 d后按照NY/T 2646—2014《水稻品种试验稻瘟病抗性鉴定与评价技术规程》记录稻瘟病发病情况。
1.4 调查与分析
根据稻瘟病菌生理小种对鉴别品种的抗感反应分别记录为抗病(R)和感病(S)。小种分类时先分群,根据小种对特特普、珍龙13、四丰43、东农363、关东51、合江18和丽江新团黑谷的感病情况,分为ZA~ZG群,如对特特普感病的属于ZA群,对特特普抗病、对珍龙13感病的属于ZB群,以此类推,对7个鉴别品种均抗病的则列为ZH群。将7个鉴别品种按照特特普、珍龙13、四丰43、东农363、关东51、合江18 和丽江新团黑谷的顺序分别给以64、32、16、8、4、2、1的固定号码,小种编号时取分群品种后面各鉴别品种中显示抗病反应的号码相加,再加1即为该小种编号(全国稻瘟病菌生理小种研究协作组,1980)。
1.5 生理小种致病力测定
将775份水稻资源进行浸种催芽,播于秧盘,3次重复,待幼苗生长至3叶期采用1.3中描述的方法分生理小种进行接种鉴定,记录苗瘟发病等级,3次重复以发病最重进行统计。高抗为0级,抗病为1级和2级,中抗为3级,感病为4级以上。根据以下公式计算致病力(邓云,2021)。
1.6 统计分析
利用Excel 2010分别对来源于不同市(县、区)的稻瘟病菌生理小种进行分析,统计安徽省各稻作区稻瘟病菌生理小种类型,分析安徽省稻瘟病菌优势种群和优势生理小种以及各稻作区优势生理小种类型。
2 结果与分析
2.1 安徽省稻瘟病菌样本来源
2019—2020年对安徽省5个稻作区21个市(县、区)的水稻稻瘟病样本进行收集和小种分离,对其中367个稻瘟病菌小种进行鉴定分类。主要包括沿淮淮北单季稻作区的凤台县、怀远县和谢家集区,病穗品种为中籼和中粳,鉴定的稻瘟病菌单孢菌株73株;江淮丘陵单、双季稻过渡区为全椒县、寿县和肥西县,病穗品种为中籼和中粳,稻瘟病菌单孢菌株33株;沿江双、单季稻作区为青阳县、无为市、宣州区、当涂县、贵池区、南陵县和怀宁县,病穗品种为早籼、中籼、中粳和晚粳,稻瘟病单孢菌株112株;大别山地单、双季稻作区为金寨县、潜山市和岳西县,病穗品种为中籼和中粳,稻瘟病菌单孢菌株56株;皖南山地单、双季稻作区为石台县、休宁县、宁国市、黄山区和歙县,病穗品种为中籼、中粳和晚粳,稻瘟病菌单孢菌株93株(表1)。

表1 安徽省主要稻作区稻瘟病样本采集信息Table 1 Collection information of rice blast samples from main rice growing areas in Anhui Province
2.2 安徽省稻瘟病菌生理小种类型
通过鉴定分类,分离获得的稻瘟病菌为7个种群25个生理小种,主要为ZA、ZB、ZC、ZE、ZF、ZG和ZH,没有ZD群小种出现,优势种群为ZB群,出现频率为58.58%,次优势种群为ZC群,出现频率为22.62%,其他种群分别为ZG群9.26%、ZH群4.90%、ZA群3.00%、ZE群0.82%和ZF群0.82%(图1)。优势生理小种为ZB13、ZB15和ZC15,出现频率分别为32.43%、17.98%和17.98%,ZG1、ZH1、ZC13、ZB29、ZB31、ZB9和ZB11生理小种亦有相对较高的出现频率,依次为9.26%、4.90%、3.54%、1.91%、1.91%、1.36%和1.36%,其他15个生理小种出现频率较低,共7.36%(图2)。

图1 安徽省主要稻作区稻瘟病菌种群出现频率Fig.1 Frequency of M. oryzae populations in main rice growing areas of Anhui Province
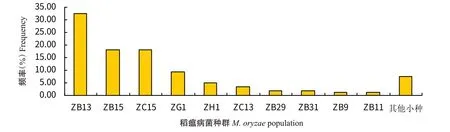
图2 安徽省主要稻作区稻瘟病菌生理小种出现频率Fig.2 Frequency of physiological races of M.oryzae in main rice growing areas of Anhui Province
优势种群ZB群中有9个生理小种,分别为ZB1、ZB5、ZB7、ZB9、ZB11、ZB13、ZB15、ZB29和ZB31,其中ZB13出现频率最高,占ZB群小种的55.35%,ZB15次之,占30.70%,ZB29和ZB31分别占3.26%,其他ZB小种共占7.43%(表2)。优势种群ZC群中有4个生理小种,分别为ZC1、ZC11、ZC13和ZC15,其中ZC15出现频率最高,占ZC群小种的79.52%,ZC13次之,占15.66%,ZC1 和ZC11 分别占1.20%和3.61%(表3)。

表2 ZB群中各小种的出现频率Table 2 Frequency of physiological races in ZB population

表3 ZC群中各小种的出现频率Table 3 Frequency of physiological races in ZC population
2.3 安徽省不同稻作区稻瘟病菌生理小种分布
安徽省地处暖温带与亚热带的过渡地带,依据地理位置、光温水土资源、耕作制度、品种类型及生产条件等,安徽水稻种植区域可划分为5个稻作区。对不同稻作区稻瘟病菌生理小种的鉴定,可为水稻品种的选育和推广布局提供参考。
对沿淮淮北单季稻作区凤台县、怀远县和谢家集区进行稻瘟病菌生理小种鉴定,凤台县鉴定出4群7个生理小种,优势小种为ZB15、ZC15和ZB13;怀远县4群9个生理小种,优势小种为ZB13和ZB9;谢家集区2群4个生理小种,优势小种为ZB13和ZG1(图3)。

图3 沿淮淮北单季稻作区稻瘟病菌小种分布Fig.3 Physiological race distribution of M.oryzae in single-season rice growing area along the Huaihe River
对江淮丘陵单、双季稻过渡区全椒县、寿县和肥西县进行稻瘟病菌生理小种鉴定,全椒县鉴定出3群4个生理小种,优势小种为ZB15;寿县3群8个生理小种,优势小种为ZB13、ZC15和ZB15;肥西县2群5个生理小种,优势小种为ZB13和ZB15(图4)。
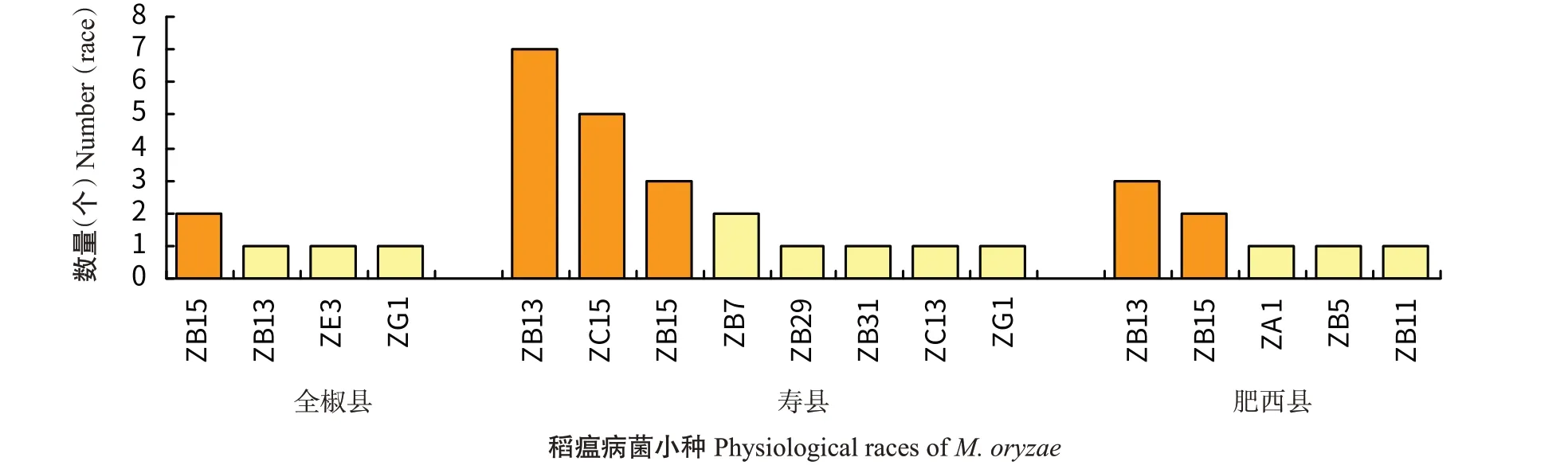
图4 江淮丘陵单、双季稻过渡区稻瘟病菌小种分布Fig.4 Physiological race distribution of M.oryzae in single-and double-season rice growing areas of the Jianghuai hilly area
对沿江双、单季稻作区青阳县、无为市、宣州区、当涂县、贵池区、南陵县和怀宁县进行稻瘟病菌生理小种鉴定,青阳县鉴定出3群4个生理小种,优势小种为ZC15;无为市4群6个生理小种,优势小种为ZB13和ZG1;宣州区4群10个生理小种,优势小种为ZB13、ZB15和ZC15;当涂县2群4个生理小种,优势小种为ZB13、ZC13和ZC15;贵池区2群3个生理小种,优势小种为ZB15和ZC15;南陵县4群9个生理小种,优势小种为ZC15、ZB13和ZB15;怀宁县1群1个生理小种,优势小种为ZB13(图5)。
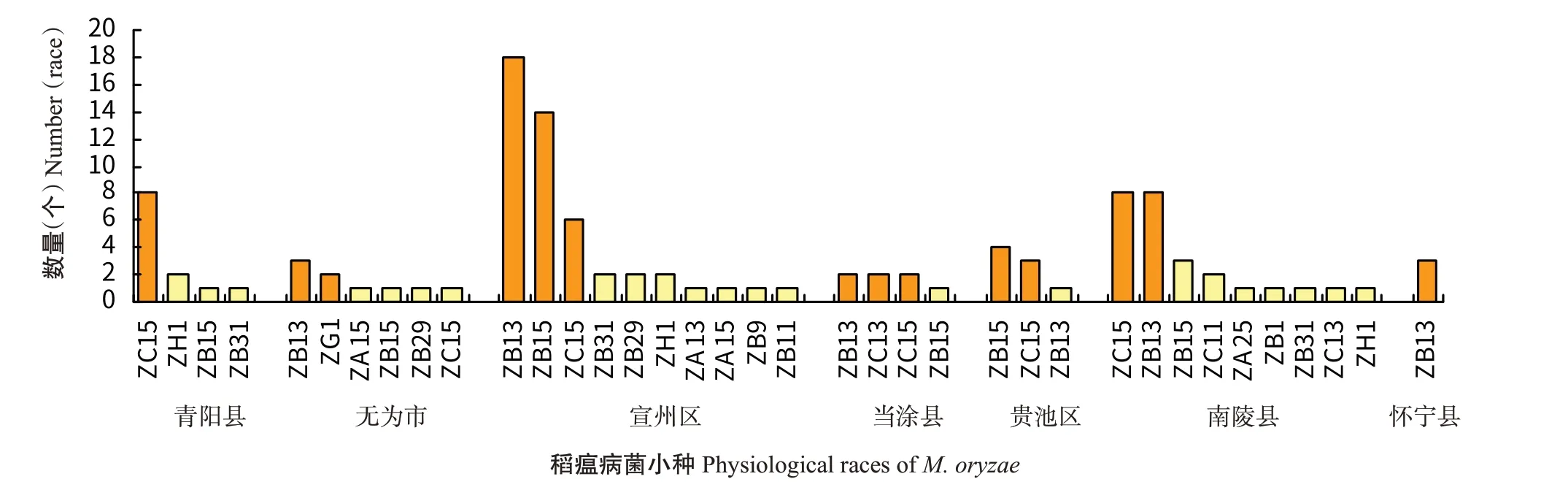
图5 沿江双、单季稻作区稻瘟病菌小种分布Fig.5 Physiological race distribution of M.oryzae in double-and single-season rice growing areas along Yangtze River
对大别山地单、双季稻作区金寨县、潜山市和岳西县进行稻瘟病菌生理小种鉴定,金寨县鉴定出3群5个生理小种,优势小种为ZB13和ZG1;潜山市2群4个生理小种,优势小种为ZB13和ZB15;岳西县3群5个生理小种,优势小种为ZB13和ZH1(图6)。
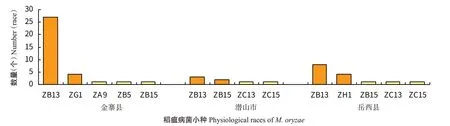
图6 大别山地单、双季稻作区稻瘟病菌小种分布Fig.6 Physiological race distribution of M.oryzae in single-and double-season rice growing areas of Dabie Mountains
对皖南山地单、双季稻作区石台县、休宁县、宁国市、黄山区和歙县进行稻瘟病菌生理小种鉴定,石台县鉴定出6群12个生理小种,优势小种为ZB15、ZB13和ZC15;休宁县5群7个生理小种,优势小种为ZC15、ZG1、ZB15和ZH1;宁国市5群6个生理小种,优势小种为ZB13和ZG1;黄山区2群3个生理小种,优势小种为ZB13、ZC13和ZC15;歙县1群1个生理小种,优势小种为ZC15(图7)。
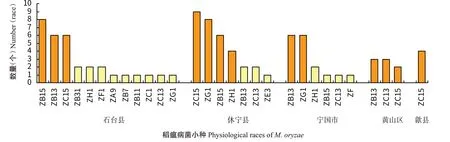
图7 皖南山地单、双季稻作区稻瘟病菌小种分布Fig.7 Physiological race distribution of M.oryzae in single-and double-season rice growing areas of mountains in south Anhui
2.4 稻瘟病菌优势生理小种致病力分析及抗苗瘟资源筛选结果
为进一步弄清安徽省优势生理小种的致病力,选取休宁县和金寨县的ZB13生理小种、休宁县和潜山县的ZB15生理小种以及休宁县和南陵县的ZC15生理小种分别对775份水稻种质资源进行苗瘟接种鉴定,并分析各小种的致病力。结果(图8)显示,休宁县和金寨县ZB13 的致病力分别为59.68%和62.06%,平均为60.87%;休宁县和潜山县ZB15的致病力分别为61.16%和64.84%,平均为63.00%;休宁县和南陵县ZC15的致病力分别为71.29%和66.26%,平均为68.78%。
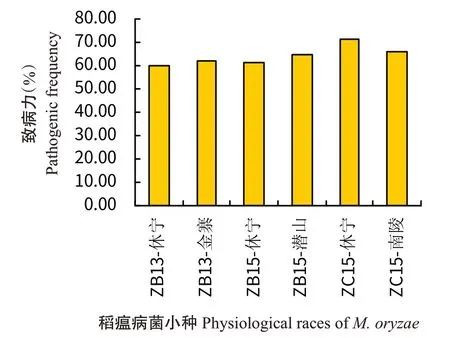
图8 优势生理小种致病力分析Fig.8 Pathogenicity analysis of dominant physiological races
通过对775份水稻种质资源鉴定筛选,共获得10份对安徽省稻瘟病菌优势生理小种表现为高抗或抗病的优异种质材料(表4),为稻瘟病抗性新基因的挖掘和育种利用提供材料基础。

表4 抗苗瘟优异种质资源筛选Table 4 Screening of excellent germplasms with seedling blast resistance
3 讨论
陈莉等(2007)对2002年收集的安徽省稻瘟病菌生理小种鉴定发现,ZA~ZH等8个种群均有分布,优势种群为ZB群,出现频率为77.2%,优势生理小种为ZB13和ZB15,出现频率分别为43.2%和25.0%。本研究通过对2019—2020年收集的安徽省主要稻作区的367个稻瘟病菌进行分离鉴定,共鉴定出ZA、ZB、ZC、ZE、ZF、ZG和ZH等7个种群,未检测到ZD群,其中优势和次优势种群为ZB和ZC群,优势生理小种为ZB13、ZB15和ZC15。研究发现,近年来安徽省稻瘟病菌种群结构与陈莉等(2007)的研究结果相比发生了显著变化,优势种群ZB群出现频率由77.2%下降至58.58%,ZC种群由3.5%上升至22.62%;优势生理小种ZB13出现频率由43.2%下降至32.43%,ZB15由25.0%下降至17.98%,优势生理小种ZC15显著上升,其出现频率增加至17.98%;其他种群的出现频率亦发生较大变化,ZA群由5.2%降至3.00%,ZD群由1.7%降至0,ZE群由1.7%降至0.82%,ZF群由1.7%降至0.82%,ZG群由5.2%上升至9.26%,ZH群由3.5%上升至4.90%;生理小种中ZG1、ZH1和ZC13亦有较高的出现频率,分别为9.26%、4.90%和3.54%。此次收集的稻瘟病病穗包括早中晚稻、籼粳糯稻、常规稻和杂交稻等,几乎涵盖了安徽省所有水稻种植类型,同时病穗来源地包括安徽省5个稻作区的主要市(县、区),稻瘟病菌菌株的收集在品种和地理位置上均较全面。本研究对安徽省不同市(县、区)的稻瘟病菌生理小种进行分离和鉴定,得出安徽省主要稻作区稻瘟病菌生理小种结构分布。研究结果表明稻瘟病菌在相同稻作区的不同市(县、区)差异明显,为安徽省水稻抗性品种的推广布局提供参考。
通过与各省稻瘟病菌优势生理小种比较发现,安徽省稻瘟病菌优势生理小种与广东省极为相似,均 为ZB13、ZB15 和ZC15,广 东 省 优 势 生 理 小 种ZB13、ZB15 和ZC15 的 出 现 频 率 分 别 为32.47%、18.05%和12.14%(钟宝玉等,2018);与湖北随州地区和湖南桃江病圃均存在相似的优势小种,为ZB15,湖北随州地区优势生理小种ZB15出现频率为60.53%,湖南桃江病圃优势生理小种为ZB15 和ZB1,出现频率分别为39.6%和15.1%(李杰等,2021;吴尧等,2021);与广西区存在相似的优势小种,为ZB13,广西区优势生理小种为ZB9和ZB13,出现频率均为19.01%(陈小林等,2017)。与江苏、山东、福建、黑龙江和辽宁地区的稻瘟病菌结构差异较大。研究结果为稻瘟病菌群体结构分析和水稻品种的推广布局提供依据和指导。
稻瘟病菌优势生理小种的致病力对水稻种质资源挖掘和品种选育至关重要,辛威等(2016)在研究黑龙江省2013—2014年稻瘟病菌优势生理小种的致病力时发现,优势小种ZD5和ZD7均具有较强的致病力;福建省优势生理小种ZB5和ZA5在不同地区亦具有较强的致病力(邓云,2021)。本研究选取不同市(县、区)的ZB13、ZB15和ZC15为优势生理小种代表进行致病力分析,结果表明这些优势小种的致病力均在50.00%以上,暗示安徽省稻瘟病菌优势生理小种不仅分布广泛且具有较强的致病力。对长江中下游地区粳稻稻瘟病菌基因型与优势混合生理小种的研究发现Pi5和Pia基因对抗性贡献较大,基因组合Pi5+Pia、Pi5+Pia+Pik+Pikm和Pita+Pik+Pikm+Pb1具有重要的应用价值(周坤能等,2022)。陈晴晴等(2022)研究结果表明长江中下游区试籼稻品种稻瘟病抗性最佳基因组合为Pi9+Pikh、Pi9+Pik+Pikh和Pi9+Pik+Pi1+Pikh。这些结果为长江中下游水稻品种选育及安徽省水稻品种推广布局提供指导。
4 结论
通过对2019—2020年安徽省不同稻作区稻瘟病菌进行鉴定,结果表明安徽省稻瘟病菌优势种群为ZB和ZC群,优势生理小种为ZB13、ZB15和ZC15,同时分析安徽省5个稻作区不同市(县、区)稻瘟病菌生理小种类型,明确其优势种群和生理小种的结构分布,为稻瘟病抗性育种和品种推广布局提供参考。
猜你喜欢
古今农业(2022年2期)2022-08-15 01:40:30
作物学报(2022年6期)2022-04-08 01:26:44
中央民族大学学报(自然科学版)(2017年4期)2017-10-22 11:31:26
植物保护(2017年4期)2017-08-09 01:18:38
中央民族大学学报(自然科学版)(2017年4期)2017-06-11 07:17:36
中国马铃薯(2017年1期)2017-03-02 09:15:51
河南农业(2016年6期)2016-11-26 07:06:46
作物学报(2016年10期)2016-10-19 04:14:09
现代农业(2016年5期)2016-02-28 18:42:37
中国茶叶(2016年4期)2016-01-18 13:29:30
