
黄淮大豆主产区大豆胞囊线虫生理小种分布调查
2016-10-19 04:14云王金社李海朝魏荷李金英武永康雷晨芳
作物学报 2016年10期
练 云王金社李海朝魏 荷李金英武永康雷晨芳
张 辉1王树峰1郭建秋2李月霞2李志辉3靳巧玲3徐淑霞4
张志民4杨彩云5于会勇5耿 臻6舒文涛6卢为国1,*
1河南省农业科学院经济作物研究所 / 国家大豆改良中心郑州分中心 / 农业部黄淮海油料作物重点实验室, 河南郑州 450002;2洛阳农林科学院, 河南洛阳 471023;3漯河市农业科学院, 河南漯河 462300;4安阳市农业科学院, 河南安阳 455000;5濮阳市农业科学院, 河南濮阳 457000;6周口市农业科学院, 河南周口 466001
黄淮大豆主产区大豆胞囊线虫生理小种分布调查
练 云1,**王金社1,**李海朝1魏 荷1李金英1武永康1雷晨芳1
张 辉1王树峰1郭建秋2李月霞2李志辉3靳巧玲3徐淑霞4
张志民4杨彩云5于会勇5耿 臻6舒文涛6卢为国1,*
1河南省农业科学院经济作物研究所 / 国家大豆改良中心郑州分中心 / 农业部黄淮海油料作物重点实验室, 河南郑州 450002;2洛阳农林科学院, 河南洛阳 471023;3漯河市农业科学院, 河南漯河 462300;4安阳市农业科学院, 河南安阳 455000;5濮阳市农业科学院, 河南濮阳 457000;6周口市农业科学院, 河南周口 466001
大豆胞囊线虫病(SCN)在黄淮地区普遍发生, 调查病原小种分布情况, 确定优势小种对抗病育种有重要意义。2012—2015年, 取样调查黄淮地区6个省份土样, 利用Riggs模式鉴定生理小种, 绘制黄淮地区SCN生理小种分布图, 并与文献报道结果对比, 探讨黄淮地区 SCN生理小种类型及其分布规律。结果表明, 该病害在黄淮大豆主产区均有分布, 在采集受SCN感染的322份土样中, 112份被鉴定出生理小种类型, 包括1号、2号、3号、4号、5号、6号和11号小种。其中, 57份为2号小种, 占样本总体的50.9%; 26份土样为5号小种, 占23.2%; 11份土样为4号小种, 占9.8%, 1号、3号、6号和11号小种分别占总体的4.5%、5.4%、4.5%和1.8%。依据不同生理小种在各省发生频率由高到低的顺序, 河南分布5号、2号、3号、11号小种; 河北分布2号、5号、6号、3号、4号小种; 安徽分布2号、5号、6号、3号小种; 山西分布2号、4号、5号、1号、3号、11号小种; 山东分布2号、3号、5号、1号、6号小种; 江苏分布2号、5号、1号小种。以上结果表明, 2号小种是目前黄淮地区的优势小种, 其次是5号小种, 致病力最强的4号小种主要分布在山西省。在黄淮地区, 抗线虫育种目标应以抗2号生理小种为主, 兼抗5号小种, 部分地区应以兼抗2号和4号小种为主。在黄淮地区3号、6号和11号小种是新发现的小种。与2001—2003年调查结果比较, 黄淮地区大豆胞囊线虫生理小种组成及分布有一定的改变。
大豆; 大豆胞囊线虫; 黄淮地区; 生理小种
大豆胞囊线虫病(soybean cyst nematode, SCN)是一种世界性的大豆病害, 给大豆生产造成严重损失[1-2], 培育抗病品种是抵御 SCN最经济有效的办法。调查大豆主产区生理小种类型及分布情况, 明确育种目标, 是培育抗病品种的先决条件。
Niblack等[3]于1998—1999年, 在美国密苏里州采取392份土壤样品, 在183份土样的鉴定中, 3号生理小种是优势小种, 其次是 1号、2号生理小种,这 3个小种占所鉴定出生理小种样本总体的 86%;Willson等[4]于 1992—1995年在美国俄亥俄州调查667块大豆田发现, 91块大豆田受到大豆胞囊线虫感染, 在其鉴定的 33份样品中发现 8个生理小种,优势小种是3号小种, 其次是1号、2号、4号、5号、6号、10号和14号小种; 2005年, Mitchum等[5]在密苏里州 8个生态区125个田块采集土样, 检测到 49%的样品受到大豆胞囊线虫感染, 他们依据HG模式, 确定了其中 45个样品的生理小种类型。上述研究均是在同一地区采集大量样本(超过100个)来详细描述小种的分布规律。
黄淮海流域包括黄河中下游地区、海河流域和淮河以北地区, 是中国大豆的主产区, 在我国大豆生产中占有重要地位[6-7]。何艳琴等[6]和卢为国等[8]研究表明, 大豆胞囊线虫在河南、河北、北京、安徽、山西和山东均有分布。通过局部取样调查发现,黄淮地区大豆胞囊线虫生理小种分布比较复杂, 存在多个生理小种, 包括张磊等[9]在安徽蒙城鉴定的5号小种, 在安徽砀山、濉溪、亳州鉴定的4号小种;陈品三等[10]在山东半岛西部的胶县、潍坊、高密、昌邑以及河南的开封、滑县、温县等地鉴定的 7号小种; 刘汉起等[11]于1983年在山东垦利、山西太谷和安徽砀山鉴定的4号小种; 李卫东等[12]、邢邯等[13]分别在在河南省鉴定的1号、2号、4号、5号小种和山东省鉴定的1号、2号、4号小种等; 李莹等[14]在太原鉴定的 4号小种; 王衍桐等[15]在北京发现的14号小种和卢为国等[8]在河南发现的9号小种。综上, 已鉴定的包括1号、2号、4号、5号、7号、9号、14号小种。上述研究多基于对个别省份或个别地点土样的鉴定, 缺乏系统调查分析, 仅卢为国等[8]于2001—2003年对黄淮地区的32个土样进行了生理小种鉴定, 并结合前人研究, 绘制了一张黄淮大豆胞囊线虫生理小种分布图。
与国外有关研究相比, 国内在大豆胞囊线虫生理小种分布调查研究方面, 取样点较为稀疏, 难以反映局部地区存在的不同大豆胞囊线虫生理小种交叉分布的现象[8]。因此, 较为密集的采样调查才能充分反映该地区大豆胞囊线虫小种分布规律; 其次,对黄淮地区大豆胞囊线虫生理小种的调查研究[8]已过去了十年之久, 在此期间, 该地区种植业结构发生了较大变化, 玉米、花生等同季作物的轮作, 是否对土壤中大豆胞囊线虫群体结构有影响是本研究关注的另一问题。
本研究拟在前人及本课题组前期调查的基础上,在黄淮海各省大豆种植区域密集采取土样, 调查该地区大豆胞囊线虫生理小种分布规律与地域分布动态, 明确优势小种, 为SCN抗性育种提供依据。
1 材料与方法
1.1 供试材料
Riggs模式鉴别寄主[16](Lee、Peking、Pickett、PI88788和 PI90763)由河南省农业科学院经济作物研究所大豆课题组保存, 2012年提纯, 2013年在河南省农业科学院现代农业科技试验示范基地扩繁,同年10月收获的成熟籽粒用于本试验。2012—2013年于大豆生长期, 在河南、河北、安徽北部、山西、山东、江苏北部的大豆产区调查大豆胞囊线虫分布情况, 在有大豆胞囊线虫感染的地块取土样322份,每份土样20~30 kg。
1.2 线虫繁殖
将采集到的土样根据土壤粘性不同, 按照病土和无菌细沙以 3.0∶0.5~1.0的比例均匀混合制备成试验病土, 将子叶出土期(萌发约3~5 d)的Lee幼苗移栽至病土中, 每杯 1株(杯子规格为 6 cm ×12 cm)。于每年5月中旬至9月中下旬, 在河南省农业科学院现代农业科技试验示范基地大豆胞囊线虫培养专用塑料大棚里进行试验。培养条件[17]为自然温度; 每天浇水 2次, 浇水前/后土壤含水量 4%~7%/ 15%~18%; 自然湿度。约25 d后, 将大豆植株从病土中倒出, 采用淘洗-过筛法从根系中分离胞囊,用研钵轻轻研磨, 把胞囊破碎, 以看不到白色胞囊为准, 然后将释放出来的卵冲洗入烧杯内, 用清水将虫卵悬浮备用, 取 10 µL, 在显微镜下计数虫卵密度。
1.3 接种方法
将壤土和沙子以2∶1混合后在150℃烘干2 h,装在塑料杯中, 每45杯装在一个67 cm ×45 cm的塑料筐中。将4个鉴别寄主及Lee在蛭石中育苗, 在子叶出土期(萌发 3~5 d), 挑选生长一致的幼苗, 移栽到塑料杯中, 每杯1株, 每个鉴别寄主播种9杯。待长出新根(移栽3~5 d)后, 接种制备好的虫卵悬浮液, 约每杯4000个卵。接种后25 d左右、胞囊处于显囊盛期时, 轻轻挤压塑料杯, 待杯子中的土松动之后, 将植株拔出, 计数根部胞囊数目。
1.4 胞囊计数
利用淘洗-过筛法从根系中分离胞囊, 使其均匀分散到一块黑布上, 用普通数码相机照相, 拍摄从每颗植株上冲洗的胞囊照片。利用本课题自主研发的 PDS软件, 通过计算机扫描、分析, 自动计数每张照片上的大豆胞囊数目, 具体方法见专利2014SR060158[18], 取在各寄主上生长的胞囊平均值,并根据Riggs模式判别生理小种类型。Lee上平均胞囊数目大于 60, 认定此次试验数据有效, 否则, 将通过再次扩繁该土样中的胞囊, 重复接种 1次, 如果Lee上平均胞囊数目仍然达不到要求, 舍弃该样本。
1.5 数据分析及小种分布图绘制
用“检出频率”表示该地区某种生理小种样本数占该地区所鉴定出生理小种样本总体的比例。本文定义为, 检出频率= (属某一种生理小种的样本数目/鉴定出生理小种类型的所有样本数目)×100%。
将各采样点鉴定出的小种类型, 利用统计分析软件R的Maptools包[19]绘制生理小种分布图。
2 结果与分析
2.1 生理小种的总体分布情况
在河南、河北、安徽、山西、山东和江苏省采集大豆胞囊线虫感染的 322份土样, 大豆胞囊线虫分布情况见图1。含 210份因胞囊密度小而未鉴定出生理小种类型的样本和 112份鉴定出生理小种类型的样本, 鉴定出的小种类型包括1号、2号、3号、4号、5号、6号和11号, 各小种构成比例见表1。分析有生理小种的112份土样表明, 2号生理小种最多, 检出频率为50.9%; 其次是5号和4号小种, 检出频率分别为23.2%、9.8%。其中, 3号、6号和11号小种是首次在黄淮地区发现。河北、安徽、山东发现的6号小种及在河南、山西发现的11号小种为国内初次报道。

图1 黄淮地区大豆胞囊线虫感染地块及生理小种分布图Fig.1 Sampling places infected by SCN and race distribution of SCN in Huang-Huai Rivers Valley
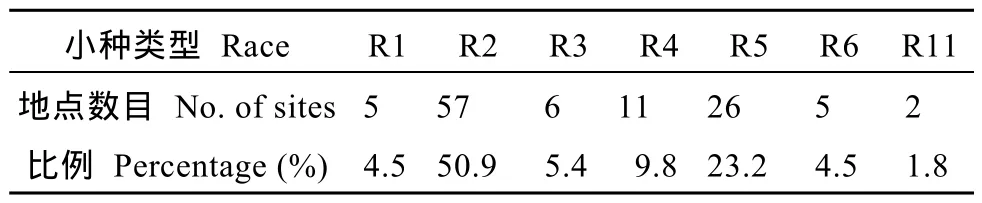
表1 黄淮地区各个生理小种所占比例Table1 Proportion of races in Huang-Huai Rivers Valley
2.2 各生理小种在黄淮大豆主产区的分布情况
从图1可以看出, 1号生理小种主要分布在山西南部、山东和江苏北部; 2号和5号生理小种在这6个省份均有分布; 3号生理小种分布在河南、河北、安徽、山西和山东; 4号生理小种主要分布在山西和河北; 6号生理小种主要分布在河北、安徽和山东;在河南和山西发现有11号生理小种。
2 号生理小种在黄淮海大豆主产区均有分布,集中分布在河北中部、南部, 江苏北部和安徽省北部, 是黄淮海地区的优势小种。
4 号生理小种集中分布在山西省中部和北部地区, 占所鉴定出4号生理小种样本的90.9%。在黄淮海其他地区, 仅在河北省邯郸市平乡县鉴定出有 4号生理小种。4号生理小种致病力最强, 抗源相对较少, 但是该小种分布集中。在山西省, 2号和4号生理小种检出频率分别是38.5%和38.5%。所以, 在山西省抗病育种应该以兼抗2号和4号生理小种为主要目标。
5 号生理小种在黄淮海各省均有分布。在河南省和安徽省分布广泛, 在黄淮海地区, 5号生理小种分布范围仅次于 2号生理小种, 是涉及黄淮海地区抗病育种工作的重点小种之一。
其他类型生理小种中, 大豆胞囊线虫 1号生理小种主要有 3片集中分布区域, 即山西南部的运城市、山东南部的枣庄市和江苏省西北部的徐州市沛县和新沂市, 检出频率为4.5%; 3号生理小种在河南东南部、山西北部及山东、安徽、江苏三省交界处有分布, 检出频率为5.4%; 6号生理小种在河北石家庄市井陉县、河北省保定市涿州市、安徽省毫州市涡阳县、安徽省阜阳市阜南县、山东省泰安市东平县有分布, 检出频率为4.5%; 11号生理小种在河南省永城市和山西省临汾市霍州市有零星分布, 检出频率为1.8%。
2.3 黄淮地区大豆主产省份大豆胞囊线虫生理小种构成及分布区域
由表2可见, 在河北、江苏、安徽、山东、山西和河南, 2号生理小种检出频率分别是 68.00%、61.50%、52.40%、45.50%、38.50%和37.50%; 5号生理小种在以上所在省份的检出频率分别是16.00%、23.10%、33.30%、18.18%、7.70%和50.00%。这 2个小种所占比例之和, 在河南最高, 达到87.50%; 在最低的山西省, 也达到了46.20%。

表2 黄淮地区各省大豆胞囊线虫不同生理小种构成Table2 Proportions of SCN races in the six provinces of Huang-Huai Rivers Valley
图2表明, 在安徽阜阳和亳州主要分布5号生理小种, 淮北和宿州一带主要分布有2号生理小种, 部分地区零星分布有3号和6号生理小种; 河北省主要分布有 2号生理小种, 部分地区兼有 5号生理小种,零星分布有3号、4号和6号生理小种; 河南省郑州、洛阳、濮阳、安阳一带大豆胞囊线虫生理小种分布较为复杂, 2号和5号生理小种交叉分布, 局部地区零星分布有3号和11号生理小种; 江苏省淮安市、宿迁市一带主要分布有2号生理小种, 在徐州市一带主要分布有 5号生理小种, 部分地区兼有 1号生理小种;在山东的菏泽、济宁、枣庄一带主要分布有2号生理小种, 局部地区零星分布有1号、3号、5号和6号小种; 山西吕梁、太原及其以北地区主要分布有4号生理小种, 兼有2号生理小种, 山西临汾、长治及其以南主要分布有2号生理小种, 局部地区零星分布有1号、3号、5号和11号生理小种。
表3表明, 在2001—2003年调查研究中, 1号生理小种是优势小种, 其次是 4号生理小种; 在2012—2015年调查研究中, 2号生理小种是优势小种, 在黄淮海大豆主产区均有分布, 检出频率是50.9%, 集中分布在河北中部、南部, 江苏北部和安徽北部。

表3 不同年份黄淮地区大豆主产省份优势小种构成的比较Table3 Composition of predominant races in the main soybean producing area of Huang-Huai Rivers Valley in different years

图2 黄淮地区6省份大豆胞囊线虫生理小种构成及主要小种分布区域Fig.2 Composition and distribution of the main SCN race in six provinces of Huang-Huai Rivers Valley
3 讨论
3.1 大豆胞囊线虫生理小种鉴别模式选择
在对大豆胞囊线虫生理小种类型研究过程中,前人依据致病力将SCN分为不同类型的生理小种,目前通用的鉴别方法有Riggs和Schmitt[16]于1988年建立的16个小种模式以及Niblack等[20]在2002年建立的“HG类型”, 利用7个鉴别寄主将大豆胞囊线虫分为 128个小种。其目的均是通过鉴别寄主区分致病力不同的线虫群体, 两者的不同之处在于HG模式将小种划分得更细。这两种划分方法各有其适宜的使用范畴, 而从抗病育种目标出发,我们认为小种不宜划分过细, 加之生理小种存在区域间更替现象, 线虫分布较为复杂, 划分过细很容易给育种带来困惑。为了便于同国内不同研究者的调查结果对比, 本研究仍然选用Riggs鉴别模式[16]探讨黄淮大豆主产区大豆胞囊线虫生理小种的分布。
3.2 黄淮地区大豆胞囊线虫生理小种分布概况
在大豆胞囊线虫生理小种鉴定研究中, 无论采用Riggs鉴别模式还是HG模式都需要足够的线虫群体接种于 4个或 7个鉴别寄主。有研究表明, 接种虫卵数目需限定在一定范围内, 否则鉴定结果不稳定[17]。因此, 为能繁殖出够量的胞囊, 我们采集土样时要求每个土样取20~30 kg。但由于土样采集困难, 因此国内目前已报道的研究, 多局限于对个别省份或个别地点的土样鉴定。卢为国等[8]的调查较为系统, 但采样点较为稀疏且没有标识经纬度。本研究是在其基础上, 对黄淮大豆主产区大豆胞囊线虫生理小种分布与地域分布动态做进一步调查研究,此次共采集病土 322份, 其中 112份病土鉴定出生理小种类型, 因此, 本次调查的结果比较系统全面地反映黄淮地区大豆胞囊线虫生理小种分布的概况,而且每份土样均有精确的经纬度记录, 这为今后的进一步研究奠定了基础。从本次调查结果来看, 黄淮地区大豆胞囊线虫发生普遍, 而且小种类型多,构成复杂。结合前人文献报道, 目前在黄淮地区发现的小种类型有 1号、2号、3号、4号、5号、6号、7号、9号、11号、14号等[6,8-15], 2号小种为优势小种, 3号、6号和11号小种均是首次在黄淮地区发现。
在调查中, 虽然有322个地点发现胞囊线虫, 但仅有1/3 (112份)鉴定出小种类型。其余210份土样是因为没有繁殖到足够的胞囊而未获得可靠的结果。因此, 可以认为图1中彩色点代表了田间胞囊量比较大的地点, 从侧面反映了黄淮地区田间种群密度的概况。对比采集病土的取样点和鉴定结果(图1)可见, 鉴定出的生理小种集中分布在山西省、河北省和河南黄河沿岸, 这些地区属于沙性土壤, 土质适宜大豆胞囊线虫滋生, 因此鉴定出生理小种的样本较多; 在黄淮区域种植大豆较为集中的安徽阜阳、安徽亳州、河南周口和河南商丘, 采集的病土较为密集, 但由于大豆胞囊线虫种群密度较小, 因此, 鉴定出生理小种的样本较少, 推测其原因, 主要是因为这些地区属于砂姜黑土, 土质黏性较大,不利于大豆胞囊线虫繁殖、扩散。
3.3 大豆胞囊线虫生理小种的动态变化规律
在相对较长的时间周期内, 优势生理小种构成是在变化的。从各生理小种在黄淮大豆主产区的分布看, 在2001—2003年调查研究中, 1号生理小种是优势小种, 检出频率是40.0%, 集中分布在山东济南及以东地区, 河南北部的安阳、滑县、延津以及河北南部的邯郸及河南漯河、周口及安徽阜阳一带[8];在2012—2015年调查研究中, 2号生理小种是优势小种, 在黄淮海大豆主产区均有分布, 检出频率是50.9%, 集中分布在河北中部、南部, 江苏北部和安徽北部。从黄淮地区大豆主产省份生理小种构成及分布看(表3), 在2001—2003年调查研究中, 河南、山东的优势小种均是1号和4号小种, 而在2012—2015年调查研究中, 河南的优势小种变为2号和5号小种,山东为2号小种; 在先后2次调查研究中, 安徽、江苏的优势小种分别从4号、5号和1号、5号均演变为优势小种为2号和5号小种; 河北的优势小种由1号、2号转变成2号小种; 山西省优势小种由4号演变成2号和4号小种。
在一定时期内, 大豆胞囊线虫生理小种分布是相对稳定的。本研究在2012—2015年, 选择河南省农业科学院现代农业科技试验示范基地连续4年监测, 鉴定结果均为2号生理小种, 此研究结果与卢为国等[8]研究结果相同。由于研究条件受限, 卢为国等[8]在2001—2003年的调查研究没有标识经纬度, 所以很多地点无法一一对应, 但从2001—2003年和2012—2015年这两个阶段的调查研究结果看, 大豆胞囊线虫生理小种在一定时期内是稳定的。
大豆胞囊线虫生理小种变化的可能原因, 首先,在黄淮海生态区, 大豆的同期作物有玉米、花生和棉花等, 玉米种植面积在2014年比2002年增加313万公顷, 而大豆种植面积同比减少50万公顷(http://zzys.agri.gov.cn/nongqing.aspx), 这些作物的轮作及种植结构的改变, 对大豆胞囊线虫群体结构有一定的影响; 其次, 农作物育种周期是十年, 在轮回的十年中, 各地抗性育种目标会随着生产中遇到的病虫害危害程度不同, 进行不同程度的调整, 这也给胞囊线虫的生存带来一定的选择压, 对抗性作物敏感的小种逐渐被淘汰, 因此大豆胞囊线虫的群体结构为了适应新的生存环境也不断地在变化。其变化周期和更详尽的原因有待进一步研究, 这也是我们下一阶段研究的重点。
3.4 生理小种的区域交叉分布
从上述结果看, 黄淮地区大豆胞囊线虫生理小种构成复杂, 而且小种分布区域互相交叉。为进一步探讨不同小种间交叉分布规律, 我们在较小的范围内密集取样, 试图明确各小种的分布边界。从河北石家庄市调查取样的方圆1209 km2看(图3), 在一定的区域内, 小种分布没有绝对界限。我们在鹿泉市李村镇3502农场采集到了2号土样, 在其东南方向的藁城市堤上村和西南方向的井陉县吴家窑乡苗峪均采集到了5号土样, 而在紧临苗峪南部3.28 km但仍属于吴家窑乡的一块大豆田采集到了6号土样;而从保定市调查取样的方圆5121 km2看, 生理小种分布在一定区域内是相对稳定的, 我们在定州市东亭镇、保定市博野县、保定市高阳县南蒲口乡北蒲村、保定市清苑县孙村乡大候乡、保定市顺平县南清醒村采集到的均是2号土样。以上研究结果表明,由于大豆胞囊线虫是一种土传的定居性内寄生线虫, 其传播速度及传播范围具有一定的局限性, 因此具有在局部地区优势小种分布相对稳定、部分地区存在其他小种的特点。这与卢为国等[8]的研究结果一致。
3.5 黄淮大豆胞囊线虫生理小种分布特点及其对育种工作的启示
本研究结果表明, 在黄淮地区大豆胞囊线虫分布有一定的区域性。从各省大豆胞囊线虫分布情况看, 在河北省、江苏省主要分布2号小种, 在河南和安徽主要分布2号和5号小种, 山西以2号和4号为主; 山东兼有2号、3号和5号小种。
国家大豆品种区域试验在黄淮地区设3个片区[6]。黄淮海夏大豆北组包括河北宁晋、河北石家庄、山东德州、山西北部等地, 主要分布有2号生理小种,其次是4号和5号生理小种; 中组包括山东潍坊、河北邯郸、山西南部、河南濮阳、河南洛阳等地, 主要分布有2号和5号生理小种, 兼1号和6号生理小种;南组包括江苏北部、安徽北部、河南郑州、河南驻马店、河南周口、山东济宁、山东菏泽等地, 主要分布2号生理小种, 其次是5号生理小种, 兼1号和6号生理小种。整个黄淮地区以抗2号小种为主要育种目标, 北组需要兼抗4号小种, 中组和南组需要兼抗5号小种。

图3 河北省大豆胞囊线虫生理小种分布图Fig.3 Distribution of SCN race in Hebei province
SCN的抗性是受多个位点控制的数量性状, Rhg位点之间相互作用[21-26], 多篇文献报道[27-28]: 大豆的第18染色体(G连锁群)上的rhg1位点与抗病基因紧密连锁, 控制着对多个大豆胞囊线虫生理小种的抗性, 因此培育多抗品种是可能的。鉴于生理小种的变化, 在抗线虫育种中, 我们更需要培育兼抗2个或2个以上生理小种的大豆品种。
4 结论
黄淮海地区大豆胞囊线虫分布广泛, 约分布于70%的地区。黄淮地区存在1号、2号、3号、4号、5号、6号和11号生理小种, 其中 2号小种为该地区的优势小种, 其次是5号生理小种, 而4号生理小种主要集中分布在山西, 3号、6号和11号小种为新发现的小种。大豆胞囊线虫在多数地区的分布呈连续状, 组成较为稳定, 有区域性分布的特点, 但在少数的局部地区没有明显规律, 在夹缝区域有其他小种存在的特点。整个黄淮海地区大豆育种的目标应以抗2号生理小种为主, 部分地区应以兼抗2号和5号生理小种为主, 而在山西应以兼抗2号和抗4号生理小种为主。
致谢: 本文在调查方案制定、论文写作等方面得到南京农业大学盖钧镒院士的悉心指导, 在此深表谢意。
References
[1] Wrather J A, Koenning S R.Estimates of disease effects on soybean yields in the United States 2003 to 2005.J Nematol, 2006,38: 173-180
[2] Niblack T L, Tylka G L, Riggs R D.Nematode pathogens of soybean.Wilcox J R.Soybeans: Improvement, Production, and Uses.Madison, WI, USA: American Society of Agronomy, Crop Science Society of America, and Soil Science Society of America,2004.pp 821-851
[3] Niblack T L, Wrather J A, Heinz R D, Donald P A.Distribution and virulence phenotypes of Heterodera glycines in Missouri.Plant Dis, 2003, 87: 929-932
[4] Willson H R, Riedel R M, Eisley J B, Young C E, Jasinski J R,Wheeler T A, Kauffman P H, Pierson P E, Stuart M C.Distribution and virulence phenotypes of Heterodera glycines in Missouri.J Nematol, 1996, 28: 599-603
[5] Mitchum M G, Wrather J A, Heinz R D, Shannon J G, Danekas G.Variability in distribution and virulence phenotypes of Heterodera glycinesin Missouri during 2005.Plant Dis, 2007, 91:1473-1476
[6] 何艳琴, 闫晓燕, 吴存祥, 杨中路, 年海.中国大豆新品种动态.北京: 中国农业科学技术出版社, 2013 He Y Q, Yan X Y, Wu C X, Yang Z L, Nian H.The dynamic of new soybean variety in China.Beijing: China Agricultural Science and Technology Press, 2013
[7] 练云, 卢为国.大豆抗 SCN机制及抗病相关基因研究进展.中国油料作物学报, 2013, 35: 727-732 Lian Y, Lu W G.Advances on resistance mechanism and gene to SCN in soybean.Chin J Oil Crop Sci, 2013, 35: 727-732 (in Chinese with English abstract)
[8] 卢为国, 盖钧镒, 李卫东.黄淮地区大豆胞囊线虫(Heterodera glycines Ichinohe)生理小种的抽样调查与研究.中国农业科学, 2006, 39: 306-312 Lu W G, Gai J Y, Li W D.Sampling survey and identification of races of soybean cyst nematode (Heterodera glycines Ichinohe) in Huang-Huai Valleys.Sci Agric Sin, 2006, 39:306-312 (in Chinese with English abstract)
[9] 张磊.安徽淮北地区大豆胞囊线虫生理小种研究初报.大豆科学, 1988, 7: 251-254 Zhang L.Preliminary research on races of soybean cyst nematode (Heterodera glycines Ichinohe) in Huaibei area of Anhui province.Soybean Sci, 1988, 7: 251-254
[10] 陈品三, 张东生, 陈森玉.大豆胞囊线虫7号生理小种的研究初报.中国农业科学, 1987, 20(2): 94 Chen P S, Zhang D S, Chen S Y.First report on a new physiological race (race 7) of soybean cyst nematode (Heterodera glycines).Sci Agric Sin, 1987, 20(2): 94 (in Chinese with English abstract)
[11] 刘汉起, 商邵刚, 甄鸿杰, 霍虹, 吴和礼, 姚振纯, 李秀兰.大豆胞囊线虫生理小种研究初报.大豆科学, 1985, 4:131-136 Liu H Q, Shang S G, Zhen H J, Huo H, Wu H L, Yao Z C, Li X L.Study on physiological races of soybean cyst nematode (Heterodera glycines).Soybean Sci, 1985, 4: 131-136 (in Chinese with English abstract)
[12] 李卫东, 贺春林, 田保明.河南省大豆胞囊线虫分布及生理小种鉴定.华北农学报, 1991, 6(增刊1): 111-114 Li W D, He C L, Tian B M.Sampling survey and identification of races of soybean cystnematode (Heterodera glycines Ichinohe) in the province of Henan.Acta Agric Boreali-Sin,1991, 6(S1): 111-114 (in Chinese with English abstract)
[13] 邢邯, 赵经荣, 战明奎, 李佩廷, 盖钧镒.山东省大豆胞囊线虫生理小种的鉴定.中国油料作物学报, 1997, 19(4): 61-65 Xing H, Zhao J R, Zhan M K, Li P T, Gai J Y.Identification of races of soybean cystnematode (Heterodera glycines Ichinohe)from Shandong province.Oil Crops China, 1997, 19(4): 61-65 (in Chinese with English abstract)
[14] 李莹, 王志, 焦广音, 常汝镇.中国大豆遗传资源对大豆胞囊线虫 4号生理小种的抗性鉴定研究.中国农业科学, 1991,24(5): 64-69 Li Y, Wang Z, Jiao G Y, Chang R Z.Studies on resistance of soybean germplasm resources to race 4 of soybean cyst nematode.Sci Agric Sin, 1991, 24(5): 64-69 (in Chinese with English abstract)
[15] 王衍桐, 彭德良, 陈受宜.灰布支黑豆对大豆孢囊线虫(Heterodera glycines) 14号小种的抗性遗传.遗传学报, 2000,27: 146-150 Wang Y T, Peng D L, Chen S Y.Inheritance of resistance to Heterodera glycines race 14 in Huibuzhi black bean.Acta Genet Sin, 2000, 27: 146-150 (in Chinese with English abstract)
[16] Riggs R D, Schmitt D P.Complete characterization of the race scheme for Heterodera glycines.J Nemato, 1988, 20: 392-395
[17] 练云, 魏荷, 王金社, 雷晨芳, 李海朝, 武永康, 卢为国.影响大豆胞囊线虫生理小种鉴定因素探讨.分子植物育种,2015, 13: 1259-1264 Lian Y, Wei H, Wang J S, Lei C F, Li H C, Wu Y K, Lu W G.A study on the factors that influence the race-identification of soybean cyst nematode.Mol Plant Breed, 2015, 13:1259-1264 (in Chinese with English abstract)
[18] 王金社, 卢为国, 李金英, 练云, 魏荷, 李海朝, 雷晨芳.植物病虫害表型数据采集系统.专利号: 2014SR060158,中国, 2014 Wang J S, Lu W G, Li J Y, Lian Y, Wei H, Li H C, Lei C F.The data acquisition system on the phenotype of plant diseases and insect pests.Patent number: 2014SR060158,China, 2014
[19] Team R C.R: a language and environment for statistical computing.R Found Statist Comput, 2014
[20] Niblack T L, Arelli P R, Noel G R, Opperman C H, Orf J H,Schmitt D P, Shannon J G, Tylka G L.A revised classification scheme for genetically diverse populations of Heterodera glycines.J Nematol, 2002, 34: 279-288
[21] Afzal A J, Srour A, Saini N, Hemmati N, El Shemy H A,Lightfoot D A.Recombination suppression at the dominant Rhg1/Rfs2 locus underlying soybean resistance to the cyst nematode.Theor Appl Genet, 2011, 124: 1027-1039
[22] Yuan C P, Li Y H, Liu Z X, Guan R X, Chang R Z, Qiu L J.DNA sequence polymorphism of the Rhg4 candidate gene conferring resistance to soybean cyst nematode in Chinese domesticated and wild soybeans.Mol Breed, 2012, 30: 1155-1162
[23] Glover K D, Wang D, Arelli P R, Carlson S R, Cianzio S R,Diers B W.Near isogenic lines confirm a soybean cyst nematode resistance gene from PI 88788 on linkage group.J Crop Sci, 2004, 44: 936-941
[24] Vuong T D, Sleper D A, Shannon J G, Nguyen H T.Novel quantitative trait loci for broad-based resistance to soybean cyst nematode (Heterodera glycines Ichinohe) in soybean PI 567516C.Thoer Appl Genet, 2010, 121: 1253-1266
[25] Liu X, Liu S, Jamai A, Bendahmane A, Lightfoot D A,Mitchum M G, Meksem K.Soybean cyst nematode resistance in soybean is independent of the Rhg4 locus LRR-RLK gene.Funct Integr Genomics, 2011, 11: 539-549
[26] 卢为国, 盖钧镒, 郑永战, 李卫东.大豆遗传图谱的构建和抗胞囊线虫(Heterodera glycines Ichinohe)的QTL分析.作物学报, 2006, 32: 1272-1279 Lu W G, Gai J Y, Zheng Y Z, Li W D.Construction of a soybean genetic linkage map and mapping QTLs resistant to soybean cyst nematode (Heterodera glycines Ichinohe).Acta Agron Sin, 2006,32: 1272-1279 (in Chinese with English abstract)
[27] Concibido V C, Diers B W, Arelli P R.A decade of QTL mapping for cyst nematode resistance in soybean.Crop Sci,2004, 44: 1121-1131
[28] Afzal A J, Natarajan A, Saini N, Iqbal M J, Geisler M, El Shemy H A, Mungur R, Willmitzer L, Lightfoot D A.The nematode resistance allele at the rhg1 locus alters the proteome and primary metabolism of soybean roots.Plant Physiol, 2009, 151: 1264-1280
Race Distribution of Soybean Cyst Nematode in the Main Soybean Producing Area of Huang-Huai Rivers Valley
LIAN Yun1,**, WANG Jin-She1,**, LI Hai-Chao1, WEI He1, LI Jin-Ying1, WU Yong-Kang1, LEI Chen-Fang1,ZHANG Hui1, WANG Shu-Feng1, GUO Jian-Qiu2, LI Yue-Xia2, LI Zhi-Hui3, JIN Qiao-Ling3, XU Shu-Xia4,ZHANG Zhi-Min4, YANG Cai-Yun5, YU Hui-Yong5, GENG Zhen6, SHU Wen-Tao6, and LU Wei-Guo1,*
1Zhengzhou Subcenter of National Soybean Improvement Center / Key Laboratory of Oil Crops in Huang-Huai Valleys of Ministry of Agriculture / Institute of Industrial Crops, Henan Academy of Agricultural Sciences, Zhengzhou 450002, China;2Luoyang Academy of Agriculture and Forestry Sciences, Luoyang 471023, China;3Luohe Academy of Agricultural Sciences, Luohe 462300, China;4Anyang Academy of Agricultural Sciences,Anyang 455000, China;5Puyang Academy of Agricultural Sciences, Puyang 457000, China;6Zhoukou Academy of Agricultural Sciences, Zhoukou 466001, China
Soybean cyst nematode (SCN) disease, caused by Heterodera glycines, occurs widely in the Huang-Huai Rivers Valley.Resistant cultivar is the most important solution to ocntrol this disease and reduce yield loss.Knowledge of SCN race distribution in soybean producing area is basis for SCN resistant breeding.A four-year survey from 2012 to 2015 was carried out to understand the diversity and distribution of Heterodera glycines races in the Huang-Huai Rivers Valley.A distribution map of races was constructed based on Riggs model.A total of 322 soil samples with SCN infection were collected.Seven H.glycines races were identified in 112 (34.8%) samples from six provinces, including five races described previously.Race 2 was the leading race, accounting for 50.9% in the 112 soil samples; race 5 was next to race 2 with the ratio of 23.2%; and the remaining types were races 4, 3, 1, 6, and 11, accounting for 9.8%, 5.4%, 4.5%, 4.5%, and 1.8%, respectively.The distribution of races varied greatly among the six provinces.According to frequencies from high to low, they were races 5, 2, 3, and 11 in Henan, races 2, 5, 6, 3, and 4 in Hebei, races 2, 5, 6, and 3 in Anhui, races 2, 4, 5, 1, 3, and 11 in Shanxi, races 2, 3, 5, 1, and 6 in Shandong, and races 2, 5, and 1 in Jiangsu.These results indicate that race 2 is the predominant race in the Huang-Huai Rivers Valley, followed by race 5, and the most virulent race 4 is centered in Shanxi province.Therefore, the SCN resistant breeding in this region should be mainly targeted towards race 2 with consideration of race 5 simultaneously, and race 4 also needs attention in some areas.In this study, races 3, 6,and 11 were newly detected in the Huang-Huai Rivers Valley.We also found that the race components and their distributions varied compared to the survey conducted in the same region in 2001-2003.
Soybean; Soybean cyst nematode; Huang-Huai Rivers Valley; Race
10.3724/SP.J.1006.2016.01479
本研究由国家自然科学基金项目(31371652), 河南省农业科学院科研发展专项资金项目(20137905), 国家现代农业产业技术体系建设专项(CARS-004)和河南省科技攻关项目(162102110141)资助。
This study was supported by the National Natural Science Foundation of China (31371652), the Special Fund for Scientific Research and Development of Henan Academy of Agricultural Sciences (20137905), the China Agriculture Research System (CARS-004), and the Science and Technology Project of Henan Province (162102110141).
(Corresponding author): 卢为国, E-mail: 123bean@163.com**同等贡献(Contributed equally to this work)
联系方式: 练云, E-mail: lianyun262@126.com; 王金社, E-mail: wjs33314@126.com
Received(): 2016-03-01; Accepted(接受日期): 2016-06-20; Published online(网络出版日期): 2016-07-04.
URL: http://www.cnki.net/kcms/detail/11.1809.S.20160704.0826.008.html
猜你喜欢
环境保护与循环经济(2022年7期)2023-01-03
南方农业学报(2022年11期)2022-03-18
粮油与饲料科技(2021年2期)2021-05-12
河南建材(2020年6期)2020-10-28
作物学报(2018年11期)2018-11-06
治淮(2018年6期)2018-01-30
植物保护(2017年4期)2017-08-09
中国马铃薯(2017年1期)2017-03-02
中国茶叶(2016年4期)2016-01-18
大豆科技(2014年5期)2014-03-23
