
氨基多糖对RANKL诱导的RAW264.7向破骨细胞样细胞分化的抑制作用观察
2012-07-31 09:22梁素霞vondenHoffJW许庆安
山东医药 2012年31期
梁素霞,von den Hoff JW,陈 卓,许庆安
(1天津市口腔医院,天津 300041;2荷兰奈梅亨大学正畸与口腔生物学研究中心;3浙江大学医学院附属口腔医院;4武汉大学)
氨基多糖(GAGs)是由氨基己糖与己糖醛酸组成的二糖单元聚合而成的直链高分子化合物。多数GAGs具有黏性,且分子链中的羧基和硫酸基使分子具有强酸性(聚阴离子特性),因而又被称为“酸性黏多糖”。GAGs包括肝素(heparin)、硫酸软骨素(CS)、透明质酸(HA)等,在体内常以与蛋白质结合的状态存在,是细胞外基质的主要组成成分[1]。多项研究显示氨基多糖参与调节凝血、肿瘤生长、骨代谢等多种生物学过程[2~4]。最近有研究显示氨基多糖可调节破骨细胞分化成熟,但机制尚不清楚。2011年,我们观察了肝素、HA和CS对RANKL诱导下RAW264.7向破骨细胞样细胞分化的影响。现报告如下。
1 材料与方法
1.1 细胞系与试剂 RAW264.7细胞来源于小鼠巨噬细胞系,由荷兰奈美津正畸与口腔生物学实验室提供。α-MEM培养基以及胎牛血清(FCS)购自英国Gibco,青霉素、链霉素购于荷兰生命技术公司,RANKL购自德国 Santa Cruz,肝素、CSC、高分子量HA(分子量为25~75 KDa)和TRAP染色试剂盒均购自德国 Sigma,8-well chamber slide购于德国Greiner,抗酒石酸酸性磷酸酶(TRAP)ELISA试剂盒购自德国R&D。
1.2 实验方法
1.2.1 细胞培养及干预 小鼠RAW264.7细胞复苏,于含10%胎牛血清、100 U/mL的青霉素和100 μg/mL链霉素的 α-MEM培养基中,于37℃、5%CO2细胞培养箱中,2 min后换液,去除未贴壁细胞。将8-well chamber slides置于细胞培养箱预培养24 h。细胞分别接种于8-well chamber slide,每孔200 μL细胞悬液,细胞浓度为40 000/孔。将细胞分为四组:对照组培养液中加入RANKL 90 ng/mL,肝素组加入肝素,HA组加入HA,CSC组加入CSC,浓度梯度分别为肝素:2、10、50、100 μg/mL,HA:0.1、0.5、2.5、5 μg/mL,CSC:2、10、50、100 μg/mL。每个浓度梯度做4个复孔,于37℃、5%CO2条件下培养,第2、4、6 天各换液1 次,每次每孔换液100 μL。
1.2.2 观察项目 ①细胞分化情况:分别取第3、5、7、8天的培养板进行细胞固定。PBS漂洗,2%多聚甲醛固定细胞30 min;pH9.0、浓度为0.2 M 的TRIS缓冲液37℃孵育60 min。TRAP染色试剂用孔径0.2 μm 的滤膜过滤,孵育染色 5 ~10 min,蒸馏水漂洗;苏木素复染,封片。TRAP染色阳性细胞的胞质呈粉红色、细胞核数目≥3个认为是成熟的破骨细胞样细胞,显微镜下行细胞计数。②TRAP5b含量:收集第8天各组细胞培养液,采用ELISA法测定TRAP5b含量,严格按试剂盒说明书操作。
1.3 统计学方法 采用SPSS17.0统计软件,数据以±s表示,采用单因素方差分析(one-way ANOVA)。P≤0.05为差异有统计学意义。
2 结果
2.1 细胞分化情况 各组均在第2~3天开始出现细胞核数目≥3个的融合破骨细胞样细胞。细胞体积在不同培养天数有所不同:初期体积较小,后逐渐增大,到第7~8天出现巨大破骨细胞样细胞。细胞形态呈多样性,如椭圆形、星形、三角形,可见伪足样突触。细胞质为TRAP染色阳性,呈粉红色。第7、8天的成熟破骨细胞样细胞的胞质中空泡多见。破骨细胞样细胞计数见图1、图2、图3。分析图1~3发现,肝素的抑制作用随浓度增大而增强,具有剂量依赖性。
2.2 TRAP5b含量 见表1。
3 讨论
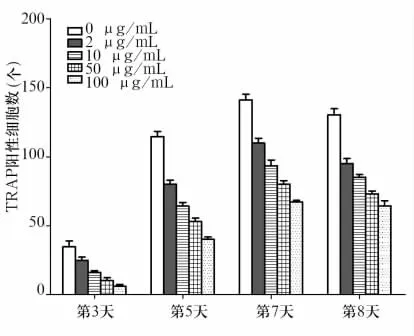
图1 肝素组与对照组不同天数TRAP阳性细胞数

图2 HA组与对照组不同天数TRAP阳性细胞数

图3 CSC组与对照组不同天数TRAP阳性细胞数
正畸过程中牙槽骨的改建包括牵张侧成骨细胞的成骨过程和压力侧破骨细胞的骨吸收过程,研究证实这一过程中有RANKL的参与[5]。成骨细胞和破骨细胞之间互相影响,达到相对稳态,此为口腔正畸的生物力学基础[6]。对成骨细胞和破骨细胞的调节尤其是前体细胞向破骨细胞分化和成熟的调控是近年来的研究热点。
越来越多的研究发现,氨基多糖作为一种重要的细胞外基质成分参与骨吸收过程,对破骨细胞的分化和成熟有诱导作用,但结果存在分歧。TRAP染色阳性及多核(≥3)特性是破骨细胞分化和成熟的标志。TRAP5b是一种酸性磷酸酶的同功酶,是破骨细胞的标志酶,既能反映破骨细胞的分化、成熟和生物活性,也能反映骨吸收的程度。
表1 各组TRAP5b含量比较(U/L,±s)

表1 各组TRAP5b含量比较(U/L,±s)
注:与对照组比较,*P <0.05;与2 μg/mL 组比较,#P <0.05;与10 μg/mL 组比较,△P <0.05;与50 μg/mL 组比较,▲P <0.05
组别 TRAP5b含量肝素组2 μg/mL 28.23 ±0.86*10 μg/mL 21.07 ±0.91*#50 μg/mL 16.96 ±0.77*#△100 μg/mL 15.18 ±0.76*#△▲HA组0.1 μg/mL 14.39 ±0.77*0.5 μg/mL 13.30 ±0.49*2.5 μg/mL 13.24 ±0.88*5.0 μg/mL 12.34 ±0.91*CSC组2 μg/mL 21.69 ±0.60*10 μg/mL 17.19 ±0.44*50 μg/mL 30.60 ±0.50*100 μg/mL 30.46 ±1.30*对照组33.86 ±1.68
Chowdhury等[7]早在1998年研究发现低浓度(5 μg/mL)肝素活化破骨细胞的活性,促进骨吸收;而 CSA、CSB、CSC 在浓度为 25 μg/mL 和 100 μg/mL时对破骨细胞分化和骨吸收作用都不明显。与之相悖的是,Ariyoshi等[8]研究发现肝素可以抑制破骨细胞的分化、成熟和功能;CSB、CSE对小鼠骨髓巨噬细胞向破骨细胞的分化具有抑制作用,但CSB对小鼠RAW264.7细胞向破骨细胞的分化无明显作用。最近的研究显示,CSA、CSC在浓度为20、100 μg/mL 时对 RAW264.7 细胞向破骨细胞样细胞的分化影响不明显,但CSB、CSD、CSE以及肝素在以上两种浓度时对破骨细胞样细胞的分化具有明显的抑制作用[9]。本研究发现不同浓度的肝素、CSC干预后破骨细胞样细胞的分化能力及活性均有所降低。相同培养天数的肝素组抑制作用随肝素浓度梯度增大而增强,具有剂量依赖性。Chang等[10]报道,HA通过调节Toll样受体家族成员4(TLR4)的功能抑制骨髓巨噬细胞向破骨细胞样细胞的分化。Ariyoshi等[11]在2005年发现低分子量 HA(相对分子质量<8 KDa)与RANKL共同作用可以上调破骨细胞样细胞的分化。本研究选取高分子量HA,发现破骨细胞样细胞的分化受到抑制,TRAP5b的含量有所减少,提示HA对破骨细胞样细胞的分化、成熟和骨吸收活性具有抑制作用。
不同的研究出现结果分歧的原因经分析可能与选择的体外培养模型有关系,如不同物种、不同细胞来源(巨噬细胞来源的纯化培养细胞或者是白血病病毒所致的肿瘤来源的细胞系)会导致结果出现差异,同时不同种类氨基多糖分子量的区别和实验浓度梯度的差异也可能会导致不同结果的出现。
综上所述,肝素、HA、CSC三种氨基多糖都对小鼠来源的RAW264.7细胞向破骨细胞样细胞的分化和成熟具有抑制作用,且肝素的抑制作用具有剂量依赖性,其具体的分子调节机制还需进一步研究。
[1]Gandhi NS,Mancera RL.The structure of glycosaminoglycans and their interactions with proteins[J].Chem Biol Drug Des,2008,72(6):455-482.
[2]Petitou M,Herault JP,Bernat A,et al.Synthesis of thrombin-inhibiting heparin mimetics without side effects[J].Nature,1999,398(6726):417-422.
[3]Hulett MD,Freeman C,Hamdorf BJ,et al.Cloning of mammalian heparanase,an important enzyme in tumor invasion and metastasis[J].Nat Med,1999,5(7):803-809.
[4]Walton KJ,Duncan JM,Deschamps P,et al.Heparin acts synergistically with interleukin-11 to induce STAT3 activation and in vitro osteoclast formation[J].Blood,2002,100(7):2530-2536.
[5]赵宁宁,林久祥,陈智滨.正畸保持期龈沟液骨保护素/核因子kappa B受体活化因子配体水平对牙槽骨改建状态的意义[J].北京大学学报,2012,44(1):108-112.
[6]Zhang P,Wu Y,Jiang Z,et al.Osteogenic response of mesenchymal stem cells to continuous mechanical strain is dependent on ERK1/2-Runx2 signaling[J].Int J Mol Med,2012,29(6):1083-1089.
[7]Chowdhury MH,Hamada C,Dempster DW.Effects of heparin on osteoclast activity[J].J Bone Miner Res,1992,7(7):771-777.
[8]Ariyoshi W,Takahashi T,Kanno T,et al.Heparin inhibits osteoclastic differentiation and function[J].J Cell Biochem,2008,103(6):1707-1717.
[9]Shinmyouzu K,Takahashi T,Ariyoshi W,et al.Dermatan sulfate inhibits osteoclast formation by binding to receptor activator of NF-kappa B ligand[J].Biochem Biophys Res Commun,2007,354(2):447-452.
[10]Chang EJ,Kim HJ,Ha J,et al.Hyaluronan inhibits osteoclast differentiation via Toll-like receptor 4[J].J Cell Sci,2006,120(Pt 1):166-176.
[11]Ariyoshi W,Takahashi T,Kanno T,et al.Mechanisms involved in enhancement of osteoclast formation and function by low molecular weight hyaluronic acid[J].J Biol Chem,2005,280(19):18967-18972.
猜你喜欢
现代畜牧科技(2021年9期)2021-10-13
中国骨质疏松杂志(2021年9期)2021-10-08
野生动物学报(2020年1期)2020-02-21
中国临床医学(2019年3期)2019-01-04
中国组织化学与细胞化学杂志(2016年3期)2016-02-27
中国骨质疏松杂志(2016年1期)2016-01-29
西安交通大学学报(医学版)(2014年5期)2014-06-23
江西理工大学学报(2013年1期)2013-03-20
郑州大学学报(理学版)(2013年3期)2013-03-11
山东医药(2012年35期)2012-07-31
