鉴别杰多霉素生物合成后修饰氧化酶JadH中参与底物结合或催化的关键残基
2012-02-10 01:20:44彭晓静季俊杰张霞范可强金玲张玉秀杨克迁
生物工程学报 2012年8期
彭晓静,季俊杰,张霞,范可强,金玲,张玉秀,杨克迁
1 中国矿业大学 (北京) 化学与环境工程学院,北京 100083
2 中国科学院微生物研究所 微生物资源前期开发国家重点实验室,北京 100101
角蒽环类聚酮抗生素是土壤中链霉菌产生的一类特殊的芳香聚酮化合物,具有化学结构和生理功能多样性。与其他芳香聚酮化合物一样,角蒽环类聚酮由Ⅱ型聚酮合酶 (Polyketide synthase,PKS) 催化合成。最小PKS (聚酮合酶α、聚酮合酶b、酰基载体蛋白) 催化短链羧酸连续脱羧聚合,形成聚酮碳链骨架[1],然后经过聚酮还原酶、环化/芳化酶等形成一个常见的角蒽环类中间产物UWM6[2]。随后的聚酮后修饰作用,如氧化还原、甲基化、糖基化等,对终产物的结构多样性起决定性作用,同时引入的基团也对产物的生物活性产生重要的影响。因此,通过对聚酮后修饰酶的研究,阐明后修饰作用的机理,应用于合成生物学中,将有助于理性合成新的“非天然”的聚酮化合物。杰多霉素 (Jadomycin,JD)是委内瑞拉链霉菌 Streptomyces venezuelae ISP5230经热激、乙醇处理或噬菌体感染产生的非典型角蒽环类抗生素[3-4],具有抗革兰氏阳性菌、酵母、及广谱的抗肿瘤活性[5-6],有重要的应用价值。研究表明杰多霉素可以通过抑制极光激酶B的活性,引发细胞凋亡[6],同时还有杀伤耐甲氧西林金黄色葡萄球菌[7]和造成 DNA损伤的活性[8]。
和其他角蒽环类聚酮化合物一样,杰多霉素也是经过对UWM6的羟化、脱水、氧化、骨架重排和糖基化等后修饰作用得到。已有的研究表明,FAD依赖的单加氧酶 JadH催化中间产物2,3-dehydro-UWM6 (DHU) 的 C12位羟化和4a,12b位脱水反应,得到 CR1,后者在空气中可以自发氧化生成另一个中间产物dehydrorabelomycin (DHR),因此JadH是一个羟化/脱水双功能酶[9]。氨基酸序列比对显示,JadH与参与角蒽环聚酮合成的FAD和NADPH依赖的单加氧酶PgaE、CabE和UrdE等有高的序列一致性。JadH与PgaE同源性为59%。PgaE参与 gaudimycin生物合成过程中两步连续的氧依赖的后修饰反应:中间产物DHU在C12和C12b位的羟化[10-11]。JadH、PgaE和CabE等与多种FAD依赖的芳香羟化酶 (FAD-dependent aromatic hydroxylases,FAH) 表现出一定的序列相似性,表明它们可能具有相同的反应机理。许多 FAD依赖的芳香羟化酶的结构和反应机制已经得到深入研究,其中存在着复杂的构象变化[12]。通过分析对羟基苯甲酸羟化酶 (p-hydroxybenzoate hydroxylase,PHBH) 的晶体结构 (1PBE) 和定点突变数据,推测在反应过程中,PHBH可能经历 3种不同构象的转化[13-14]:与底物结合的PHBH处于“in”构象;底物电离触发酶的构象变化,转为“out”构象,从而可以与NADPH结合并使FAD还原得到FADH2;释放NADP+后PHBH恢复“in”构象,FADH2被 O2氧化得到过氧化中间体,并进一步羟化底物对羟基苯甲酸,此时的“in”构象保证了活性中间体不与溶剂接触而失活;随后产物被释放,酶转为“open”构象,可以再次结合底物,开始下一轮循环。几个关键氨基酸残基的突变可以影响 PHBH的构象转换,干扰酶催化过程,其中R220Q和A45V可以稳定“open”构象[15],减缓底物结合速率;A45G可以稳定“in”构象[16],阻断反应过程。芳香羟化酶PgaE和CabE与PHBH的三维结构相似。P282S突变使PgaE活性提高[17]。JadH可能存在与PHBH和PgaE相似的反应历程和构象变化。
本研究通过构建JadH结合底物DHU的三维结构模型和氨基酸序列比对,用定点突变和体外酶学催化实验分析可能影响JadH活性的关键氨基酸,从而为进一步了解JadH结合底物的方式以及催化反应的机理提供依据。
1 材料与方法
1.1 材料
1.1.1 菌株和质粒
含jadFGHK基因的质粒pJV69A (+)、质粒pET30a (+)、E. coli BL21 (DE3) 菌株为本实验室构建或保存。
1.1.2 酶和主要试剂
KOD_Plus DNA聚合酶购自TOYOBO公司。T4 DNA连接酶和 DNA限制性内切酶购自TaKaRa (Shiga,Japan) 公司。Ni-NTA Superflow购自GE公司。化学试剂FAD和NADPH均购自Sigma-Al-drich公司。PCR扩增引物和DNA测序由上海英骏生物技术有限公司完成。DHU、DHR和CR1均由本实验室制备。
1.2 方法
DNA分子操作、感受态制备和转化均遵循《分子克隆实验指南》[18]。BLAST系列软件(NCBI) 用来从数据库中搜索相关基因或蛋白序列。多重序列比对利用Clustal X完成。蛋白的建模工作利用SWISS-MODEL Web服务器完成。蛋白三级结构的观察和作图利用 RasMol、Swiss-Pdb Viewer和MolMol完成。
1.2.1 JadH野生型表达载体的构建
天然序列的JadH和带有His6标签的JadH催化活性几乎没有差异,为了方便后续实验蛋白的纯化,构建表达载体pET30a-jadH-nHis,用来表达带有His标签的JadH蛋白。以pJV69A (+) 为模板,使用正向引物 (5¢-GTGGGTACC GTGAC CACCACCCCG-3¢) (下划线表示KpnⅠ的酶切位点) 和反向引物 (5¢-GGAATTC ACCGGGCCGC GCCGC-3¢) (下划线表示 EcoRⅠ的酶切位点),PCR得到含有完整jadH基因的片段。用KpnⅠ和EcoRⅠ双酶切jadH片段和pET30a (+)。将酶切得到的载体和目的片段连接,连接产物转化E. coli BL21 (DE3),对得到的克隆进行DNA序列测序,筛选得到构建正确的pET30a-jadH-nHis。
1.2.2 JadH突变体表达质粒的构建
以构建好的野生型 JadH的表达载体pET30a-jadH-nHis为模板,设计均含有突变位点的、部分互补的突变引物进行PCR。在PCR产物中加入1 μL DpnI酶,37 ℃酶切1 h,酶切产物转化E. coli BL21 (DE3),对得到的克隆进行DNA序列测序,筛选得到构建正确的JadH突变体表达质粒。
1.2.3 JadH野生型和突变体的表达与纯化
带有野生型和突变型 JadH表达质粒的E. coli BL21 (DE3) 在含50 mg/L硫酸卡那霉素的Luria-Bertani (LB) 培养基中37 ℃、220 r/min培养至OD600为0.4~0.6,加入IPTG至0.5 mmol/L,16 ℃、220 r/min诱导表达10 h。离心收集菌体。重悬于50 mmol/L Tris-HCl缓冲液 (pH 8.0),超声破碎后离心取上清。按Ni-NTA Superflow说明纯化JadH野生型及其突变体。将纯化得到的蛋白于30 kDa的超滤管中脱盐浓缩,重复3次。
1.2.4 蛋白浓度测定
蛋白浓度用Bradford法[19]测定。
1.2.5 体外酶学反应
将纯化得到的酶 (0.2 mmol;JadH-N-His) 加到反应混合液中 (总体积为 200 μL,含有50 mmol/L Tris-HCl 缓冲液,pH 8.0,4 μmol/L FAD,125 μmol/L NADPH和100 μmol/L DHU),32 ℃反应60 s,加入1 μL 1 mol/L的HCl终止反应。HPLC检测反应产物。
1.2.6 HPLC检测分析
反应产物CR1和DHR的定量检测使用C18反相HPLC (Shimadzu Prominence HPLC system)。洗脱速度为 1 mL/min,洗脱梯度为 25%乙腈+0.1%TFA到100%乙腈+0.1%TFA。底物DHU,产物CR1和DHR在266 nm和320 nm处检测。
1.2.7 相对活性分析
由于JadH的真正催化产物CR1在酶促反应体系中会自发氧化转化为 DHR,因此在计算JadH的催化活性时,要将产物DHR折合成CR1的量。根据对CR1自发氧化过程的HPLC分析,确定DHR与CR1的峰面积比值为1.4625∶1。因此可按下面公式计算折合产物量:
折合产物量=产物CR1峰面积+产物DHR峰面积/1.4625。
突变型JadH酶的相对活性根据折合产物量计算:
相对活性=突变体酶的折合产物量´100/野生型酶的折合产物量。
2 结果
2.1 JadH结构模型的构建和分析
以 CabE的晶体结构 (2QA1) 为模型构建JadH的三维结构模型,并使用PSI-Dock程序[20]构建JadH与DHU复合物的结构模型。结构模型分析显示,底物DHU结合于JadH的一个疏水性口袋之中,并与R50、G51、L52、F100、R221、 I223、P295、G297、G298等残基形成疏水作用和氢键,这可能是JadH与底物DHU识别和结合的结构基础。氨基酸序列比对 (图 1) 显示,PHBH的R220、A45以及PgaE的P282在JadH中保守,分别为R221、G51和P295。推测,JadH中的这些位点也可能影响JadH的构象变化和催化作用。
2.2 野生型和突变型JadH的异源表达与纯化
选择JadH活性中心内的9个氨基酸 (R50、G51、L52、G53、F100、R221、I223、P295和G298),构建了14个定点突变体 (表1),测序筛选阳性克隆。在E. coli BL21 (DE3) 中异源表达野生型JadH及其突变体,确定JadH野生型及其突变株的蛋白表达条件,SDS-PAGE显示所有JadH突变体均得到高效表达。JadH突变体蛋白用镍离子亲和层析纯化,超滤脱盐浓缩,SDS-PAGE显示蛋白纯度>95% (图2)。
2.3 HPLC测定JadH野生型及其突变体的相对活性
JadH是一个 FAD依赖的羟化/脱水双功能酶,催化DHU C12位的羟化和4a,12b位的脱水,生成具有蒽酚结构的CR1,CR1在空气中自发氧化成DHR[9]。由于CR1的自发氧化在酶促反应体系中无法避免,需要将生成的 DHR换算成CR1,合并计算确定JadH的活性。用HPLC检测野生型及突变型JadH体外酶学反应的直接产物CR1以及CR1在空气中自发氧化产物DHR的量,以野生型 JadH的催化活性为 100%,计算突变型 JadH的相对活性。结果显示突变型JadH的活性均显著低于野生型 (表1)。
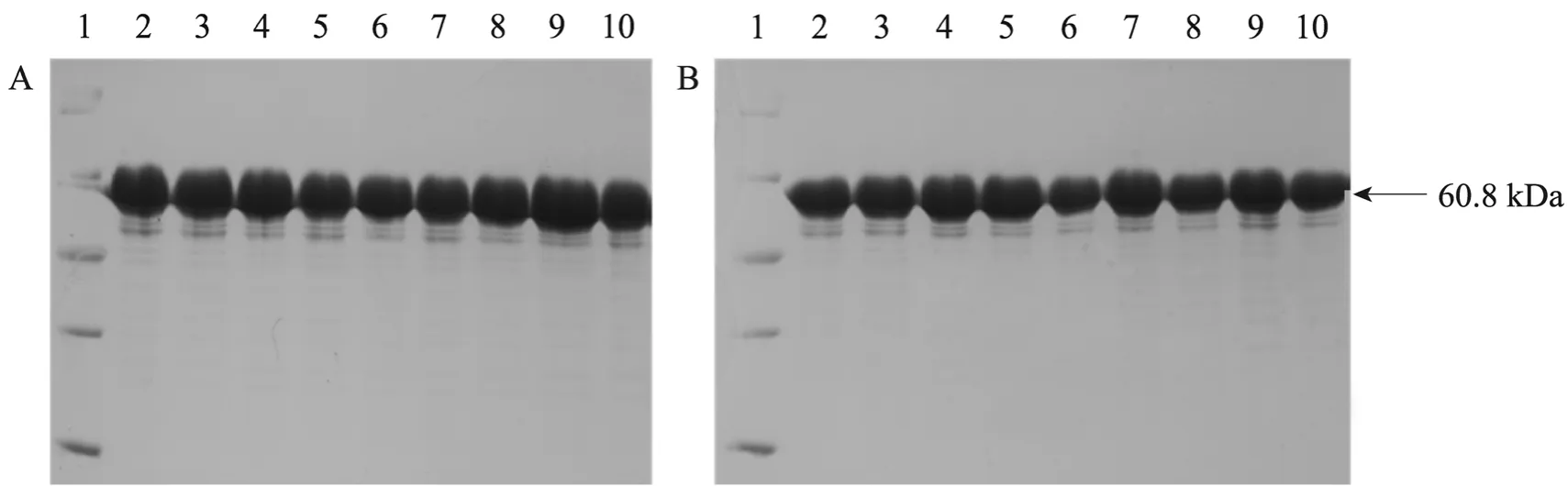
图2 纯化后JadH及其突变体蛋白的SDS-PAGE分析Fig. 2 SDS-PAGE analysis of purified wild type JadH and mutants. (A) 1: low molecular weight protein marker; 2: wild type JadH; 3: L52T; 4: L52K; 5: L52D; 6: G298K; 7: G298L; 8: G51V; 9: F100A; 10: wild type JadH. (B) 1: low molecular weight protein marker; 2: wild type JadH; 3: I223A; 4: R221Q; 5: R221L; 6: P295S; 7: G53D; 8: G53K; 9: R50L; 10: wild type JadH.
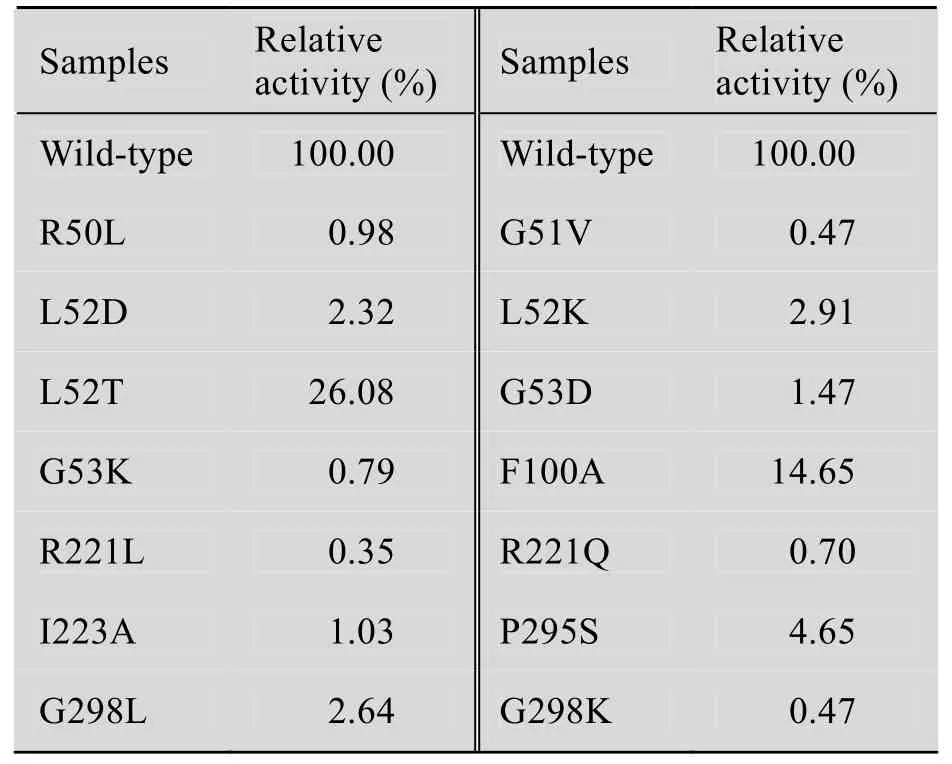
表1 JadH突变株的相对活性Table 1 The relative catalytic activities of JadH mutants
3 讨论
迄今为止,还没有关于JadH蛋白晶体结构的报道,我们依据JadH同源蛋白CabE的晶体结构构建了JadH及其与DHU复合物的结构模型(图 3)。该结构模型与 CabE的晶体结构非常类似。通过分析结构模型并与其他羟化酶和氧化酶进行序列比较,推测JadH中9个氨基酸 (R50、 G51、L52、G53、F100、R221、I223、P295和G298) 可能参与底物识别、构象转换或参与维持FAD所在区域内合适的疏水环境。我们通过定点突变得到JadH的突变体后进行体外酶学反应,通过HPLC检测产物,以JadH野生型为对照,分析JadH突变体的相对活性,对这些残基的功能进行了验证。
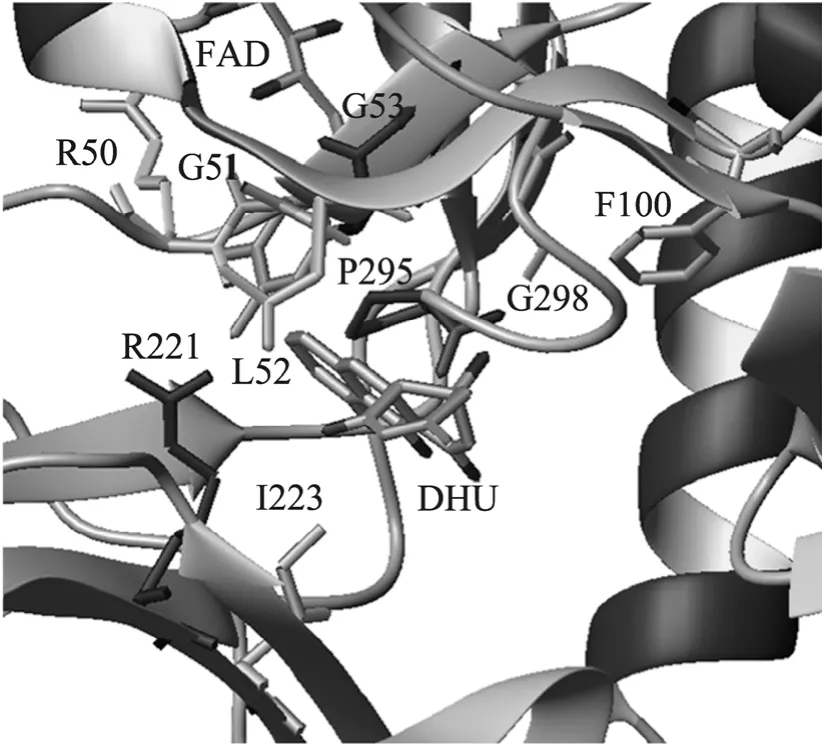
图3 JadH底物结合口袋内的关键氨基酸Fig. 3 The key residues in the substrate-binding pocket of JadH.
所得突变体的活性均远低于野生型,其中L52T与 F100A 的相对活性分别为野生型的26.08%和14.65%;L52D、L52K、I223A、G298L和P295S突变体只有微弱的催化活性 (相对活性1%~5%);G298K、R221L、R221Q和R50L均丧失了催化活性 (相对活性<1%)。R50是一个带正电荷的极性氨基酸,而 Leu为非极性氨基酸,极性的变化导致突变株R50L基本丧失催化活性。结构模型显示,I223可与底物DHU形成疏水相互作用,当将其突变为Ala时,疏水作用减弱,而且链长改变导致空间位阻效应,因此I223A只有微弱的催化活性。残基 L52和 P295可分别与底物DHU的A环和D环形成疏水相互作用,当突变为极性氨基酸Asp、Lys或Ser时,JadH与底物疏水作用减弱,识别、结合底物的能力降低,催化活性也随之降低。另外,F100侧链的苯环可能与底物DHU的C3位甲基形成较弱的疏水作用,Phe突变为Ala后,疏水作用减弱,影响底物的结合,导致活性较低。除疏水作用外,氢键也是酶与底物识别的重要结构基础。结构模型显示,G298主链上的氨基与底物的C4a位羟基可能形成很强的氢键,R221的侧链末端胍基也可能与底物的C1位羰基氧形成氢键。当将Gly突变为具有较大侧链的Leu或Lys时,产生较大的空间阻碍,干扰了氢键的形成,因此G298K丧失了催化活性,G298L也仅有微弱的活性 (2.27%)。而突变R221L和R221Q均同样也会干扰R221侧链胍基参与氢键的形成,进而影响到JadH的活性。同时,JadH的催化功能依赖FAD,推测脱水反应可能是由FAD传递质子的酸碱催化反应。疏水口袋的 FAD所在区域本身不带电荷,将L52突变为带正电的Lys或者带负电荷的Asp后,会改变底物结合区域的静电性质,可能干扰 FAD的质子传递过程,因此也会导致活性降低。同时,虽然Thr的疏水性比Leu弱,但是由于其带有一个烷基侧链,也可与底物形成疏水相互作用,因此L52T的相对活性高于L52D,为26.08%。
根据序列和结构特征,推测JadH应该具有和 FAD依赖的芳香羟化酶类似的反应机理和构象转化过程。根据PHBH的研究结果以及序列比对信息推测,JadH的G51可能参与酶的构象转化过程,G51突变成Val以后可稳定Open构象,减缓底物结合速率,从而降低酶的活性。G53距离底物的脱水位置和 FAD都比较近,将其突变成带电的Lys和Asp,会改变底物区域的静电性质,干扰 FAD在底物脱水过程中的质子传递过程,从而影响其活性,这与实验结果一致。
但令人意外的是,突变体P295S的活性远低于野生型酶,这与PgaE中对应的氨基酸P282的结果截然不同。在PgaE中将P282突变为Ser使PgaE活性提高,作者推测P282可能起到触发和传递构象变化的关键作用,P282突变为Ser后构象变化的能垒减弱,有利于构象转变[17]。在JadH的结构中,P295可能与DHU的D环形成疏水作用,可能涉及构象转换,并且P295可能与附近的L294、A296、V352、P268以及P85形成一个疏水的区域,保证活性中心氧化反应的顺利进行。当P282突变为Ser后可能会与T269形成氢键,而P295与底物DHU以及附近疏水性氨基酸之间的疏水作用被破坏,从而不利于反应的进行,所以造成了P295S活性的剧烈下降。
目前只有少数几种 FAD依赖型单加氧酶的催化机理研究得比较深入。例如,PHBH、PgaE和CabE已经得到晶体结构,并且分析了PHBH和PgaE活性中心的关键残基,推测了可能的反应机理[13,17,21]。通过异源表达和基因敲除等实验对褐黄癌菌素[22]、竹桃霉素[23]、乌达霉素[24]合成基因簇中的JadH同源蛋白进行了功能研究,推测它们可能催化不同中间体的羟化反应。还有一些聚酮合成途径中的黄素依赖型单加氧酶没有得到详细的功能研究,只是根据序列特征进行了功能推测,如 auricin合成基因簇中的 Aur1A和 Aur1I[25],simocyclinone合成基因簇中的SimA7、SimA8和Sim7[26],alpomycin合成基因簇中的 AlpG和 AlpF[27]等。除 PgaE的突变株H83A和P282S活性提高150%、K92Q活性与野生型相当,N289D和△377-491活性显著降低外(≤1%)[17],对这些芳香聚酮后修饰氧化酶结合或催化底物的可能关键氨基酸的功能分析鲜有报道。本研究首次构建了JadH的三维结构模型,除与PgaE中P282同源的P295外,第一次对JadH中的R50、G51、L52、G53、F100、R221、I223和G298进行了定点突变和体外酶学分析。实验结果表明这9个氨基酸残基是JadH底物结合或催化的关键氨基酸,推测:R50、L52、I223、P295、F100参与与底物 DHU之间的疏水相互作用;G298的主链氨基及 R221的侧链胍基与底物DHU形成氢键,参与底物的识别;G53和 L52影响FAD的质子传递;G51影响酶的构象转化。这为进一步研究JadH的催化机理及JadH近缘酶的功能提供了重要依据。
[1] Hopwood DA. Genetic contributions to understanding polyketide synthases. Chem Rev, 1997, 97(7): 2465−2498.
[2] Kulowski K, Wendt-Pienkowski E, Han L, et al. Functional characterization of the jadI gene as a cyclase forming angucyclinones. J Am Chem Soc, 1999, 121(9): 1786−1794.
[3] Doull JL, Singh AK, Hoare M, et al. Conditions for the production of jadomycin B by Streptomyces venezuelae ISP5230: effects of heat shock, ethanol treatment and phage infection. J Ind Microbiol, 1994, 13(2): 120−125.
[4] Doull JL, Ayer SW, Singh AK, et al. Production of a novel polyketide antibiotic, jadomycin B, by Streptomyces venezuelae following heat shock. J Antibiot, 1993, 46(5): 869−871.
[5] Zheng JT, Rix U, Zhao L, et al. Cytotoxic activities of new jadomycin derivatives. J Antibiot, 2005, 58(6): 405−408.
[6] Fu DH, Jiang W, Zheng JT, et al. Jadomycin B, an Aurora-B kinase inhibitor discovered through virtual screening. Mol Cancer Ther, 2008, 7(8): 2386−2393.
[7] Jakeman DL, Bandi S, Graham CL, et al. Antimicrobial activities of jadomycin B and structurally related analogues. Antimicrob Agents Chemother, 2009, 53(3): 1245−1247.
[8] Monro SM, Cottreau KM, Spencer C, et al. Copper-mediated nuclease activity of jadomycin B. Bioorg Med Chem, 2011, 19(11): 3357−3360.
[9] Chen YH, Fan KQ, He YZ, et al. Characterization of JadH as an FAD-and NAD(P)H-Dependent Bifunctional Hydroxylase/Dehydrase in Jadomycin Biosynthesis. Chembiochem, 2010, 11(8): 1055−1060.
[10] Kallio P, Patrikainen P, Suomela JP, et al. Flavoprotein hydroxylase PgaE catalyzes two consecutive oxygen-dependent tailoring reactions in angucycline biosynthesis. Biochemistry, 2011, 50(24): 5535−5543.
[11] Kallio P, Liu ZL, Mäntsälä P, et al. Sequential action of two flavoenzymes, PgaE and PgaM, in angucycline biosynthesis: chemoenzymatic synthesis of gaudimycin C. Chem Biol, 2008, 15(2): 157−166.
[12] Entsch B, Ballou DP, Massey V. Flavin-oxygen derivatives involved in hydroxylation by p-hydroxybenzoate hydroxylase. J Biol Chem, 1976, 251(9): 2550−2563.
[13] Entsch B, Cole LJ, Ballou DP. Protein dynamics and electrostatics in the function of p-hydroxybenzoate hydroxylase. Arch Biochem Biophys, 2005, 433(1): 297−311.
[14] Entsch B, van Berkel WJ. Structure and mechanism of para-hydroxybenzoate hydroxylase. FASEB J, 1995, 9(7): 476−483.
[15] Wang J, Ortiz-Maldonado M, Entsch B, et al. Protein and ligand dynamics in 4-hydroxybenzoate hydroxylase. Proc Natl Acad Sci USA, 2002, 99(2): 608−613.
[16] Cole LJ, Gatti DL, Entsch B, et al. Removal of a methyl group causes global changes in p-hydroxybenzoate hydroxylase. Biochemistry, 2005, 44(22): 8047−8058.
[17] Koskiniemi H, Metsä-Ketelä M, Dobritzsch D, et al. Crystal structures of two aromatic hydroxylases involved in the early tailoring steps of angucycline biosynthesis. J Mol Biol, 2007, 372(3): 633−648. [18] Sambrook J, Fritsch EF, Maniatis T. Molecular Cloning: a Laboratory Manual. 2nd ed. New York: Cold Spring Harbor Laboratory Press, 1989: 20−25.
[19] Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem, 1976, 72(1/2): 248−254.
[20] Pei J, Wang Q, Liu Z, et al. PSI-DOCK: towards highly efficient and accurate flexible ligand docking. Proteins, 2006, 62(4): 934−946.
[21] Ballou DP, Entsch B, Cole LJ. Dynamics involved in catalysis by single-component and two-component flavin-dependent aromatic hydroxylases. Biochem Biophys Res Commun, 2005, 338(1): 590−598.
[22] Kharel MK, Zhu LL, Liu T, et al. Multi-oxygenase complexes of the gilvocarcin and jadomycin biosyntheses. J Am Chem Soc, 2007, 129(13): 3780−3781.
[23] Zhu LL, Ostash B, Rix U, et al. Identification of the function of gene lndM2 encoding a bifunctional oxygenase-reductase involved in the biosynthesis of the antitumor antibiotic landomycin E by Streptomyces globisporus 1912 supports the originally assigned structure for landomycinone. J Org Chem, 2005, 70(2): 631−638.
[24] Rix U, Remsing LL, Hoffmeister D, et al. Urdamycin L: a novel metabolic shunt product that provides evidence for the role of the urdM gene in the urdamycin A biosynthetic pathway of Streptomyces fradiae TÜ 2717. Chembiochem, 2003, 4(1): 109−111.
[25] Novakova R, Bistakova J, Homerova D, et al. Cloning and characterization of a polyketide synthase gene cluster involved in biosynthesis of a proposed angucycline-like polyketide auricin in Streptomyces aureofaciens CCM 3239. Gene, 2002, 297(1/2): 197−208.
[26] Trefzer A, Pelzer S, Schimana J, et al. Biosynthetic gene cluster of simocyclinone, a natural multihybrid antibiotic. Antimicrob Agents Chemother, 2002, 46(5): 1174−1182.
[27] Pang X, Aigle B, Girardet JM, et al. Functional angucycline-like antibiotic gene cluster in the terminal inverted repeats of the Streptomyces ambofaciens linear chromosome. Antimicrob Agents Chemother, 2004, 48(2): 575−588.
猜你喜欢
云南化工(2021年6期)2021-12-21 07:30:56
科学(2020年2期)2020-08-24 07:57:00
安徽医科大学学报(2016年12期)2017-01-15 14:21:44
山东农业工程学院学报(2016年6期)2016-12-01 05:38:19
温州大学学报(自然科学版)(2016年1期)2016-10-27 14:57:48
生物技术通报(2015年1期)2015-04-10 16:15:19
山东医药(2015年40期)2015-02-28 14:28:45
应用化工(2014年7期)2014-08-09 09:20:23
无机化学学报(2014年5期)2014-02-28 17:31:40
食品科学(2013年23期)2013-03-11 18:30:11