
槐定碱抑制内毒素血症小鼠肺组织LPS模式识别受体的表达①
2012-01-23 12:04梁锦屏谢建宁王大军宁夏医科大学基础医学院病原生物学与免疫学系银川750004
中国免疫学杂志 2012年12期
梁锦屏 谢建宁 王大军 周 娅 (宁夏医科大学基础医学院病原生物学与免疫学系,银川750004)
近年来随着固有免疫识别机制的突破性进展与内毒素(主要成分脂多糖,即LPS)致病机理的深入研究,为内毒素疾病的防治提供了新的思路。LPS是革兰阴性菌的主要病原相关分子模式(Pathogen associated molecular pattern,PAMP),进入机体后,固有免疫系统主要通过脂多糖结合蛋白(LPS binding protein,LBP)、CD14(Cluster of differentiation 14)、Toll样受体4(Toll like receptor 4,TLR4)等模式识别受体(Pattern recognition receptors,PRRs)识别 LPS,激活TLR4炎症通路[1],促使内源性介质释放和炎症因子的过度分泌,造成全身组织器官损伤和功能紊乱。其中肺脏是最早受累器官之一,可引起急性肺损伤(Acute lung injury,ALI)、甚至急性呼吸窘迫综合征(Acute respiratory distress syndrome,ARDS)[2]。
槐定碱(Sophoridine)是苦豆子等中草药的天然提取物,具有抗肿瘤、保护心肌、影响中枢神经系统等多种药理活性[3]。本课题组既往研究发现槐定碱可减轻内毒素血症小鼠肺组织的病理损害[4]。为具体探讨槐定碱抗内毒素作用的药理机制,本研究从LPS激活的炎症信号转导通路入手,动态观察槐定碱对LPS识别受体LBP、CD14、TLR4的影响,为槐定碱防治内毒素性肺损伤提供基础资料。
1 材料与方法
1.1主要试剂与仪器 槐定碱(批号:960324,含量98%以上)购自宁夏药物研究所,其小鼠腹腔注射LD50为 58.0 mg/kg;LPS(E.coli 055∶B5,批号:63H4010)购自Sigma公司;RT-PCR试剂盒购于美国Promega公司;DNA marker购自北京博奥森生物技术有限公司;BCA蛋白浓度测定试剂盒,江苏碧云天生物技术有限公司;兔抗小鼠TLR4多克隆抗体,美国Santa Cruz公司;辣根酶标记山羊抗兔IgG,北京中杉金桥生物有限公司;小鼠TNF-α放免分析试剂盒购于北京科美东雅生物技术有限公司;PCR仪、核酸浓度测定仪、凝胶成像仪、电泳仪均为美国Bio-RAD公司;旋涡混合器(上海第一医学仪器厂XW-80型);γ放射免疫计数仪(中国科学技术大学科技实业总公司GC-1200)。
1.2动物分组 6~8周龄 BALB/c小鼠,18~22 g,雌雄各半,宁夏医科大学动物实验中心提供(合格证号:SCXK(宁)2005-2001)。BALB/c小鼠试验前自由摄取食水。随机分为6组:正常对照组、内毒素血症模型组(LPS模型组)、槐定碱药物对照组(SRI组,12 mg/kg)、槐定碱干预三个剂量组[LPS+SRI组,12 mg/kg(高)、6 mg/kg(中)、3 mg/kg(低)]。每一大组再分为2、6、12、24小时4个时相点,共24个小组,每组10只。①正常对照组:尾静脉注射生理盐水0.1 ml/10 g,②内毒素血症模型组:尾静脉注射LPS 7 mg/kg,①②两组30分钟后,均腹腔注射生理盐水0.2 ml/10g;③~⑤槐定碱高、中、低剂量干预组:尾静脉注射LPS 7 mg/kg,30分钟后,分别腹腔注射槐定碱12 mg/kg、6 mg/kg、3 mg/kg;⑥槐定碱药物对照组:尾静脉注射生理盐水0.1 ml/10 g,30分钟后腹腔注射槐定碱12 mg/kg。各组在处理完毕后的2、6、12、24小时4个时相点分别取材。
1.3检测指标 ①肺脏的形态学观察:肉眼观察肺脏大体形态;取一肺叶固定、包埋、HE染色,光镜观察。②肺水含量:剥离左肺,用吸水纸吸干表面血迹,电子天平称取左肺湿重(W),然后置80℃干烤箱72小时至恒重,再称干重(D)。计算左肺湿/干重值(W/D)。③血清肿瘤坏死因子(TNF-α)含量:小鼠摘眼球取血,3 000 r/min离心10分钟,留血清,按放免试剂盒说明书操作,γ放免计数仪检测。④RT-PCR检测肺组织LBP、CD14和TLR4 mRNA表达水平:提取小鼠肺组织总 RNA,按照一步法RT-PCR试剂盒说明书进行操作。引物由北京赛百盛公司合成,序列见表1。扩增产物于1.5%琼脂糖凝胶电泳,凝胶图像分析系统成像,Bio-Image Analysis System进行半定量分析,mRNA相对含量=目的基因条带的相对光密度值 (ROD)×面积(mm2)/β-actin条带的ROD×mm2。⑤Western blot检测肺组织TLR4蛋白表达。肺组织匀浆,提取总蛋白,BCA法测蛋白浓度。加样40 μg/孔,以 10%SDS-PAGE 电泳,经转膜和封闭后,加小鼠 TLR4一抗(1∶500);4℃过夜,PBS-T洗膜,加入HRP标记的二抗(1∶2 000),室温放置2小时,洗膜,四氯-1-萘酚显色。凝胶图像分析系统成像、扫描分析,以TLR4条带面积灰度值与β-actin条带面积灰度值的比值作为TLR4蛋白相对含量。
1.4统计学分析 采用SPSS11.5软件进行统计学分析,数据以±s表示,多样本均数间比较用单因素方差分析(One-Way ANOVA),组间比较用LSD法。

表1 目的基因序列与扩增长度Tab.1 Primer sequences and PCR products(bp)
2 结果
2.1槐定碱对内毒素血症小鼠肺脏病理形态学影响。
2.1.1肉眼观察 正常对照组与槐定碱对照组小鼠肺脏外观未见异常改变,而LPS模型组小鼠在2小时时即可见肺脏充血,随时间延长病变加重,至6、12、24小时时肺脏不仅明显充血水肿,还有出血,多为出血点,有的已呈斑片状出血灶甚至胸腔内出血。槐定碱三个剂量干预组各时相点小鼠肺脏病变均较同时相点LPS模型组明显减轻,仅几只小鼠肺组织充血水肿明显,肺表面仅见少量散在浅红色渗血。
2.1.2光镜观察 正常对照组与槐定碱对照组小鼠肺组织结构正常,肺泡腔清晰,偶见肺泡间隔轻度增宽,支气管黏膜上皮完整。LPS模型组2小时时无明显变化,随时间延长,肺组织病理变化明显加重。6、12小时组可见肺脏有广泛而强烈的炎症反应,肺泡腔内有渗出物,有的肺泡出现代偿性肺气肿,严重的间质性肺炎,大量中性粒细胞及少量单核细胞浸润,毛细血管扩张及间质小静脉扩张出血,偶可见支气管上皮纤毛倒伏;24小时组损伤亦未见减轻。槐定碱三个剂量干预组12小时与24小时时相点,与同时相点LPS模型组比较肺组织炎症损伤明显减轻(图1)。
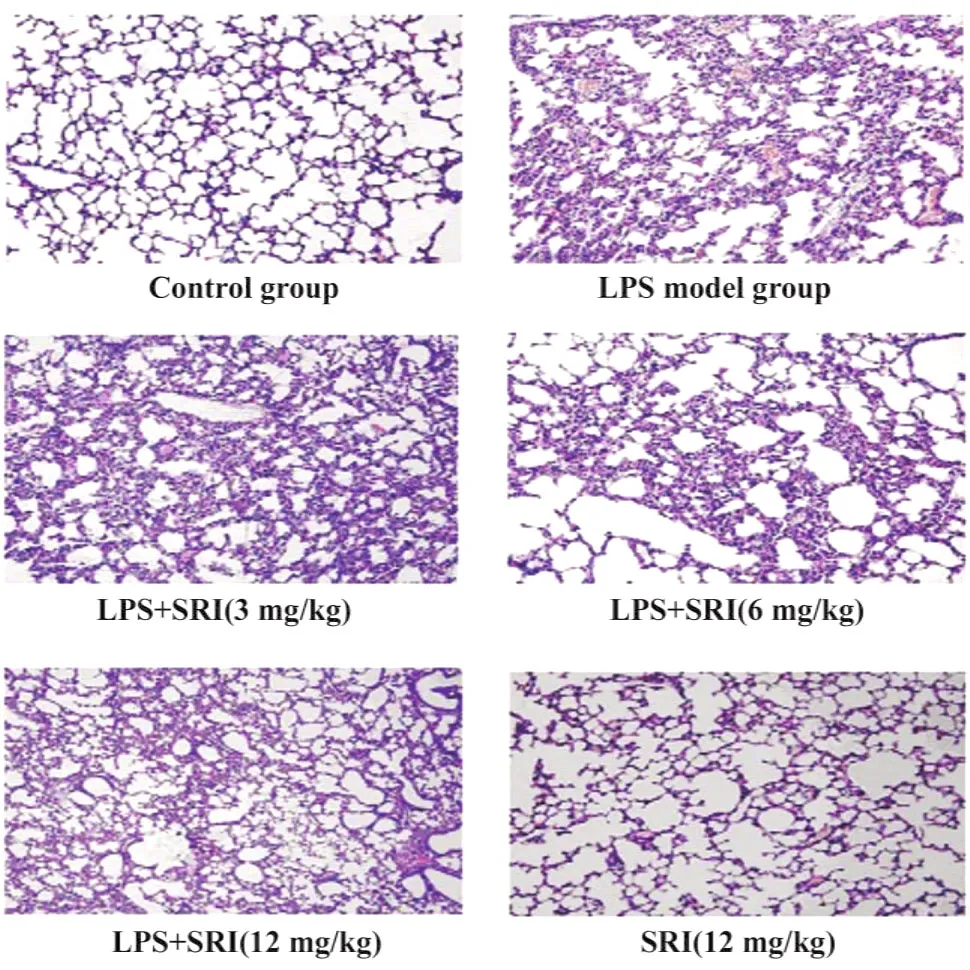
图1 光镜观察各组小鼠肺组织病理变化(24小时相点,HE×100)Fig.1 Pathological change of lung tissue of each mouse group by light microscopy(24 h,HE ×100)
2.2槐定碱对内毒素血症小鼠肺脏含水量的影响(左肺W/D值) 6、12小时时相点槐定碱高、中、低3个剂量组与同时相点LPS模型组的W/D值比较均显著降低(P<0.01或P<0.05);并与正常对照组无显著差异。槐定碱高、中、低三个剂量组同时相点之间比较,差异均无统计学意义(图2)。
2.3槐定碱对内毒素血症小鼠肺组织LBP、CD14、TLR4的mRNA表达的影响
2.3.1肺组织LBP mRNA表达的动态变化 槐定碱三个剂量干预组除2小时时相点外,其余各时相点肺组织LBP mRNA表达水平均较同时相点LPS模型组显著降低(P<0.01或P<0.05);大多数与同时相点正常对照组无统计学差异(除外槐定碱高剂量6、24 小时组,图 3、4)。
2.3.2肺组织CD14 mRNA表达的动态变化 槐定碱三个剂量干预组除2小时时相点外,其余各时相点肺组织CD14 mRNA表达水平均较LPS模型组降低(P<0.01);除槐定碱高、中剂量2、6小时组外,其余槐定碱干预组的肺组织CD14 mRNA表达水平也与同时相点正常对照组无显著差异。槐定碱高、中、低三个剂量组同时相点之间比较,差异均无统计学意义(图5、6)。
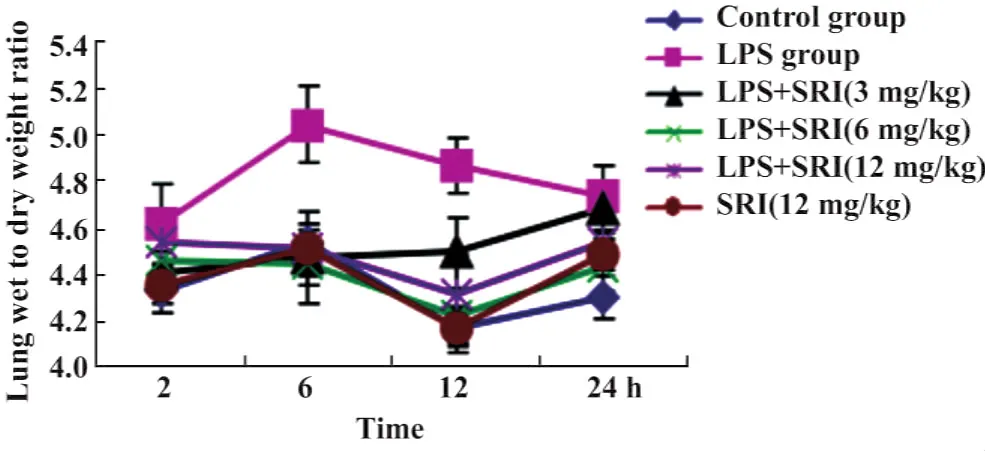
图2 槐定碱对小鼠肺脏含水量的影响Fig.2 Effect of Sophoridine on content of lung water of endotoxemia mice
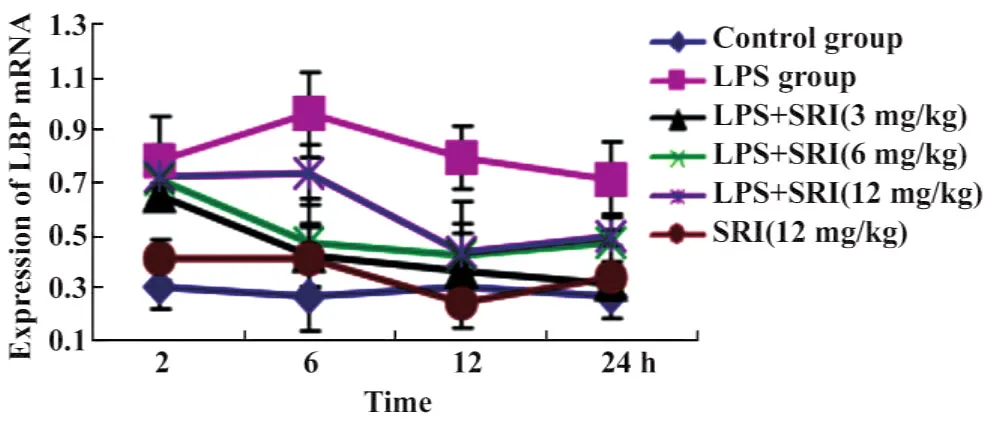
图3 槐定碱对内毒素血症小鼠肺组织LBP mRNA表达的影响Fig.3 Effect of Sophoridine on the expression of LBP mRNA in lung tissue of endotoxemia mice
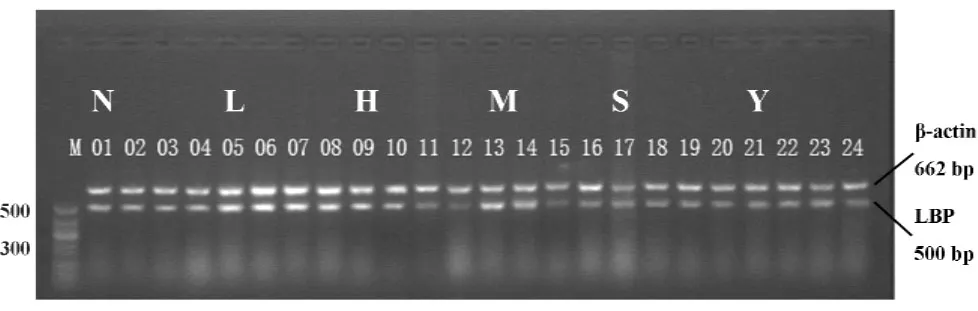
图4 RT-PCR检测各组小鼠肺组织LBP mRNA表达Fig.4 Expression of LBP mRNA in lung tissue of each mouse group by RT-PCR
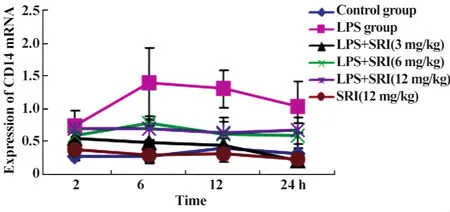
图5 槐定碱对内毒素血症小鼠肺组织CD14 mRNA表达的影响Fig.5 Effect of Sophoridine on the expression of CD14 mRNA in lung tissue of endotoxemia mice
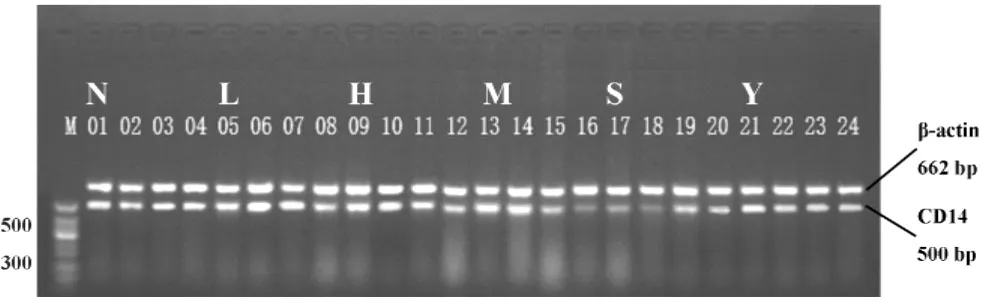
图6 RT-PCR检测各组小鼠肺组织CD14 mRNA表达Fig.6 Expression of CD14 mRNA in lung tissue of each mouse group by RT-PCR
2.3.3肺组织中TLR4 mRNA表达 槐定碱三个剂量干预组各个时相点均下调了TLR4 mRNA的表达(P<0.01或P<0.05),但仍显著高于同时相点正常对照组(P<0.01或P<0.05),直至24小时时三个剂量组TLR4 mRNA表达方降至正常对照组水平。槐定碱高、中、低三个剂量组同时相点之间比较TLR4 mRNA表达均无统计学差异(图7、8)。
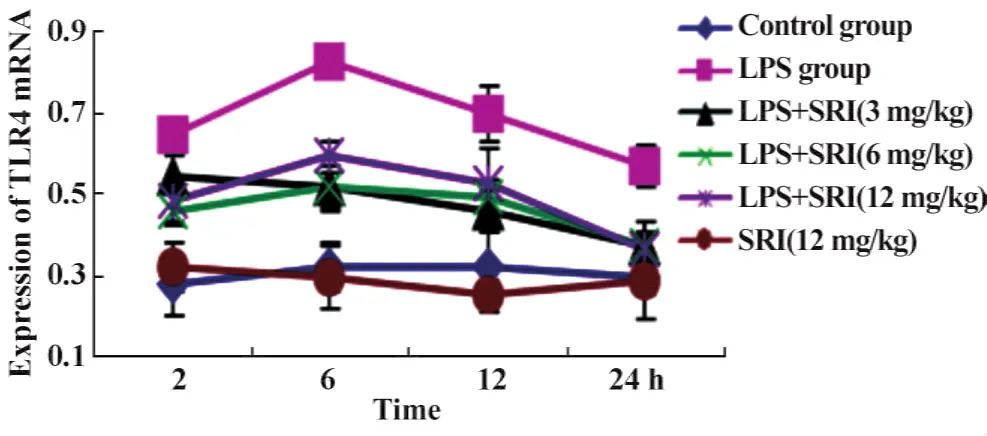
图7 槐定碱对内毒素血症小鼠肺组织TLR4 mRNA表达的影响Fig.7 Effect of Sophoridine on the expression of TLR4 mRNA in lung tissue of endotoxemia mice
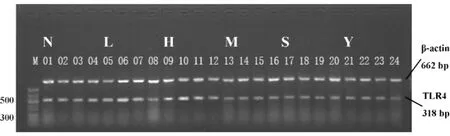
图8 RT-PCR检测各组小鼠肺组织TLR4 mRNA表达Fig.8 Expression of TLR4 mRNA in lung tissue of each mouse group by RT-PCR
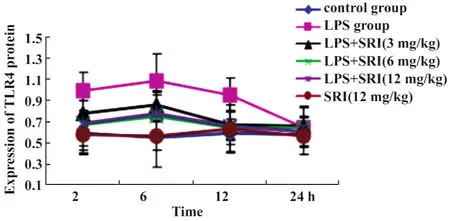
图9 槐定碱对内毒素血症小鼠肺组织TLR4蛋白表达的影响Fig.9 Effect of Sophoridine on the expression of TLR4 protein in lung tissue of endotoxemia mice
2.4槐定碱对内毒素血症小鼠肺组织TLR4蛋白表达的影响 各组小鼠肺组织中TLR4蛋白表达趋势与其mRNA相似,槐定碱三个剂量干预组2、6、12小时三个时相点TLR4蛋白表达均较同时相点LPS模型组显著降低(P<0.05);与TLR4 mRNA表达不同的是,24小时时包括 LPS模型组在内的各组TLR4蛋白表达与正常对照组比较差异均无统计学意义(均P>0.05)。槐定碱高、中、低三个剂量组同时相点之间比较TLR4蛋白表达差异均无统计学意义(图9、10)。

图10 Western blot检测各组小鼠肺组织TLR4蛋白表达(12小时时相点)Fig.10 Expression of TLR4 protein in lung tissue of each mouse group by Western blot(12 h)
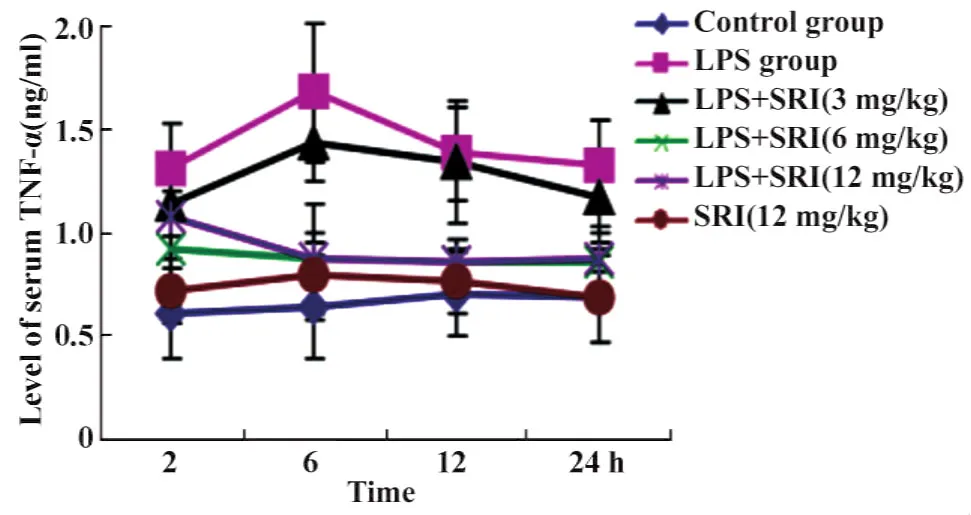
图11 槐定碱对内毒素血症小鼠血清TNF-α的影响Fig.11 Effect of Sophoridine on the level of serum TNF-α of endotoxemia mice
2.5槐定碱对内毒素血症小鼠血清肿瘤坏死因子(TNF-α)含量的影响 槐定碱高、中剂量干预组小鼠血清TNF-α含量均低于同时相点 LPS模型组(P<0.01或P<0.05),并在 6、12、24小时时均接近同时相点正常对照组;但槐定碱低剂量干预组仅6小时时TNF-α含量低于LPS模型组,各时相点均高于正常对照组(均P<0.01)。槐定碱三个剂量组同时相点之间比较,高、中剂量组血清TNF-α含量低于低剂量组,差异有统计学意义(P<0.01或P<0.05,图11)。
3 讨论
在固有免疫识别的PAMP中,LPS是研究较为深入的一种。当 G-菌感染,LPS释出后,即被LBP、CD14、TLR4等模式识别受体(PRR)识别,并启动细胞内信号转导[5]。所以说LPS识别受体是炎症反应发生的起步点。其中LBP和CD14是体内重要的脂多糖增敏系统,能极大提高多种细胞对内毒素的敏感性,对结合的LPS信号起着“放大器”的作用[6,7]。本实验中 LPS模型组小鼠在尾静脉注入LPS后2小时肺组织中 LBP、CD14、TLR4 mRNA表达均迅速上调,并于6小时达高峰,至24小时仍维持在高表达水平,各时相点表达水平均显著高于同时相点正常对照组(P<0.01);TLR4蛋白表达趋势与TLR4 mRNA基本一致,与TLR4mRNA不同的是至24小时已降至正常水平。根据已知的LPS信号转导途径,本研究制备模型时注入小鼠体内的LPS首先与血清LBP结合,再被转运至肺泡巨噬细胞表面结合CD14。此时肺泡巨噬细胞上CD14表达升高,才可结合更多的LPS-LBP复合物,而LPS结合数量的增多和结合效率的提高,又使其生物学效应得以最大程度的发挥。此后CD14传递的LPS被TLR4识别并结合,TLR4发生二聚化,募集下游的衔接蛋白,将 LPS信号向胞内转导[8-10]。本实验中TLR4在模型小鼠肺组织表达的上调显然亦是LPS作用的结果,因为要使大量被CD14结合的LPS分子有足够的跨膜受体转导信号,只有上调TLR4分子的表达,才能将LPS信号转至胞内,进一步激活下游的炎症因子通路。而大量炎症细胞因子又可促进CD14和TLR4的表达,以提高LPS信号转导的效率。这样,肺组织中LBP、CD14和TLR4的高表达与LPS所致肺脏炎症之间形成的正反馈作用,加重肺脏的局部炎症,最终发展为ALI和ARDS。
本研究用槐定碱高、中、低三个剂量干预内毒素血症肺损伤小鼠,在不同时相点都观察到槐定碱干预组肺组织 LBP、CD14 mRNA表达以及 TLR4 mRNA和蛋白的表达均较同时相点LPS模型组显著下降,在12小时和24小时时相点槐定碱三个剂量干预组上述各项指标的表达多数已与正常对照组无统计学差异。模式识别受体的下调,同时再结合小鼠一般状况显著改善,肺脏病理损伤明显减轻,肺水含量及血清TNF-α降低等疗效指标,共同表明槐定碱可明显抑制肺组织中LBP、CD14、TLR4的mRNA及TLR4蛋白表达,进而削弱这些识别受体在信号转导通路中的作用,在细胞对LPS的识别阶段即阻断其所致的病理过程,使过度的炎症反应得到控制,从而减轻因大量细胞因子和炎症介质的产生对肺组织造成的损伤,使得ALI、ARDS的发生和发展在早期得到控制。
本研究还进一步探讨了内毒素血症肺损伤小鼠血清中炎性介质TNF-α的产生与LPS识别受体的关系,结果表明槐定碱对三个识别受体表达与血清TNF-α含量表现出一致的下调作用,而且槐定碱三个剂量组抑制血清TNF-α作用有明显的剂量-效应关系,对其它指标的表达则未显示有统计学意义的剂量-效应关系。此结果一方面再次验证了槐定碱对内毒素血症小鼠肺组织的保护作用与下调TNF-α表达有关,另一方面也表明槐定碱对TNF-α分泌的抑制作用,可能来源于其对LPS激活的TLR4信号转导通路上游识别受体表达的抑制。
纵观本文研究结果,可以认为槐定碱干预使内毒素血症肺损伤小鼠肺组织LBP、CD14和TLR4的转录及TLR4蛋白表达显著下降,从而抑制LPS所致TLR4介导的胞内通路的过度活化,进而使TNF-α等炎性细胞因子的生成大大减少,而炎性细胞因子表达降低还可反过来下调LBP、CD14、TLR4受体的表达,使导致失控性炎症反应的恶性循环在此时得到遏制,阻止了肺损伤的恶性发展。这可能是槐定碱抗内毒素的信号转导机制之一。
然而TLR4信号转导通路中,不仅有不同信号分子间的直接或间接的相互影响,还存在上下游分子间的级联反应,从而形成了一个复杂的信号网络系统,调节着彼此的功能,要明确搞清槐定碱的作用机制及对其他分子的干预作用,还有待于更深入的研究。
1 Edwin S,Theo J C,Johan Kuiper.Receptors,mediators,and mechanisms involved in bacterial sepsis and septic shock[J].Clin Microbiol Rev,2003;16(3):379-414.
2 Harrison D A,Welch C A,Eddleston J M.The epidemiology of severe sepsis in England,Wales and Northern Ireland,1996 to 2004:secondary analysis of a high quality clinical database,the ICNARC Case Mix Programme Database[J].Crit Care,2006;10(2) :R42.
3 夏玉叶,王大元.槐定碱的药理作用研究进展[J].中国药学杂志,1997;32(7):392-393.
4 韩 燕,周 娅,刘 泉.苦豆子抗内毒素效应的实验研究[J].中药材,2006;29(7):1066-1068.
5 Aderem A,Ulevitch R J.Toll-like receptors in the induction of the innate immune response[J].Nature,2000;406(6797) :782-787.
6 Himanshu K ,Taro K,Shizuo A.Pathogen Recognition by Immune System[J].Int Rev Immunol,2011;30:16-34.
7 Miller S I,Ernst R K,Bader M W.LPS,TLR4 and infectious disease diversity[J].Nat Rev Microbol,2005;3(1):36-46.
8 Kawai T,Akira S.The role of patteren recognition receptors in innate immunity:Update on Toll-like receptors[J].Nat Immunol,2010;11:373-384.
9 Iwasaki A,Medzhiton R.Regulation of adaptive immunity by the innate immune system[J].Science,2010;327:291-295.
10 Park B S,Song D H,Kim H M et al.The structural basis of lipopolysaccharide recognition by the TLR4-MD-2 complex[J].Nature,2009;458:1191-1195.
猜你喜欢
中国现代医生(2022年19期)2022-11-04
中国畜禽种业(2021年10期)2021-12-02
中老年保健(2021年4期)2021-08-22
猪业科学(2021年3期)2021-05-21
医学前沿(2021年18期)2021-04-14
心肺血管病杂志(2020年5期)2021-01-14
中医眼耳鼻喉杂志(2019年3期)2019-04-13
中医眼耳鼻喉杂志(2019年2期)2019-04-13
科学中国人(2017年36期)2017-06-09
中华老年多器官疾病杂志(2016年8期)2016-05-14
