
低氧对小鼠骨骼肌离体线粒体膜通透性转换孔功能的影响
2012-01-21 03:40彭艳艳孙丽娜宋菁金露王小同
温州医科大学学报 2012年2期
彭艳艳,孙丽娜,宋菁,金露,王小同
(温州医学院附属第二医院 脑科、康复中心,浙江 温州 325027)
低氧是一种常见的应激,可引起机体结构及功能的改变,其结果随着低氧的程度、持续时间、机体反应性的不同而异。低氧可广泛影响机体的多个系统,包括神经系统、呼吸循环系统和肌肉骨骼系统等。骨骼肌作为人体最大的组织,为机体的活动提供动力。低氧可引起机体低氧应激,从而导致骨骼肌结构和功能的变化[1-3]。线粒体(mitochondria)作为机体的“能量工厂”,是机体利用氧进行呼吸功能和生成ATP的重要细胞器。低氧应激可引起线粒体功能障碍[4]。同时线粒体也与细胞凋亡和肿瘤的发生密切相关[5]。本实验旨在通过利用低氧导致小鼠骨骼肌离体线粒体低氧的模型,探讨低氧对小鼠骨骼肌离体线粒体膜通透性转换孔(mitochondrial permeability transition pore,MPTP)功能的影响。
1 材料和方法
1.1 动物与分组 SPF级C57BL/6雄性小鼠32只(由温州医学院实验动物中心提供),体质量20~28 g,7~9周龄,按随机数字表法分为低氧组和常氧组,每组16只。
1.2 试剂与仪器 线粒体分离试剂盒购自pierce公司,BCA蛋白浓度测定试剂盒购自碧云天,小鼠CytC ELISA法检测试剂盒、纯化线粒体冻存液和GENMED纯化线粒体膜通道孔比色法检测试剂盒购于上海杰美基因医药科技有限公司,超氧化物歧化酶(SOD)测试盒、谷胱甘肽(GSH)测试盒、微量丙二醛(MDA)测定试剂盒均购自南京建成科技有限公司,连二亚硫酸钠(Na2S2O4)为国产分析纯;TECNAI10透射电子显微镜(PHILIPS公司),ALLEGRA-64R台式高速冷冻离心机(美国BECKMAN公司),荧光分光光度计,MD连续波长酶标仪(美国),AVL-COMPACT3型血气分析仪(瑞士AVL公司)。
1.3 小鼠骨骼肌离体线粒体低氧模型的建立 提取线粒体后,低氧组将0.5 mL的线粒体悬液加入2 mL的密闭容器中,保持37 ℃并向内通入100%氮气及氧清除剂Na2S2O40.5 mmol·L-1致PO2测不出来,保持30 min。常氧组:线粒体悬液置于空气中,其他条件同低氧组。
1.4 取材和检测方法
1.4.1 小鼠骨骼肌线粒体的提取:小鼠直接用颈椎脱臼法处死,分离后肢股四头肌,立即放入4 ℃预冷的PBS液冲洗2次,仔细去除PBS,将组织剪碎后加800μL PBS液,玻璃匀浆器冰上研磨后1000×g,4 ℃离心3 min,弃上清液,沉淀物加入800μL LBSA/A液,漩涡振荡5 s(中速),冰上孵育最长2 min,再加入10μL B液,漩涡振荡5 s(高速),冰上孵育5 min,并且每分钟高速振荡1次,后再加入800μL C液,倒置混匀,不可振荡。700×g,4 ℃离心10 min后弃沉淀将上清转移到2 mL管中,12000×g,4 ℃离心15 min,弃上清液,保留沉淀,在沉淀中加入500μL Wash Buffer,12000×g,4 ℃离心5 min后弃上清液,得到线粒体沉淀,在沉淀中加入200μL线粒体冻存液制成线粒体悬液。整个提取过程均在冰浴中(0~4 ℃)进行,提取的线粒体冰上保存,当天检测指标。
1.4.2 线粒体蛋白含量测定:采用BCA法。
1.4.3 骨骼肌线粒体膜电位测定:取线粒体悬液50μL加入现配的1 mL JC-1染液中,再加入2 mL蒸馏水,在激发波485 nm,散发波590 nm条件下用荧光分光光度计测定反映线粒体膜电位的相对荧光单位(RFU)。JC-1染液的配制参照上海碧云天生物技术有限公司提供的试剂盒说明书操作。
1.4.4 MPTP开放状态检测:取线粒体悬液70μL加入到96孔板的对应孔里,再加入170μL GENMED缓冲液(Reagent A),混匀,即刻放进酶标仪在波长540 nm条件下读数,观察MPTP开放状态。
1.4.5 线粒体CytC测定:造模后的线粒体悬液于10000×g,4 ℃,离心10 min,取上清检测CytC具体操作见说明书。
1.4.6 SOD、GSH和MDA测定:根据蛋白定量结果取线粒体悬液适量,再用线粒体缓冲液补足至500 μL,玻璃匀浆器冰上研磨5 min破膜,10000×g 4 ℃,离心10 min, 取上清检测 SOD活性、GSH和MDA含量(具体按照南京建成提供的说明书操作)。
1.5 统计学处理方法 正态性检验采用shapirowilk检验,方差齐性采用levene检验,组间比较采用独立样本t检验。
2 结果
2.1 骨骼肌MPTP开放状态测定 以待测样本的吸光值比值来代表离体线粒体MPTP的开放状态,吸光值越低,表示MPTP开放增加。与常氧组比较,低氧组骨骼肌线粒体的吸光值比值明显降低,提示低氧组MPTP开放增强,差异有统计学意义(P<0.01)见表1。
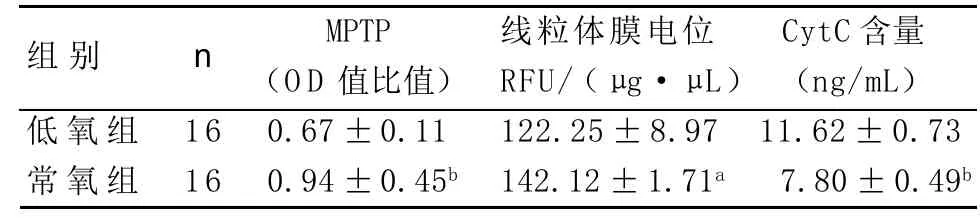
表1 线粒体膜通道孔开放、线粒体膜电位和CytC含量
2.2 线粒体膜电位测定 与常氧组相比,荧光分光光度计的定量测量结果提示低氧组线粒体膜电位明显降低,差异有统计学意义(P<0.05),见表1 2.3 线粒体释放的CytC含量测定 与常氧组相比低氧组线粒体释放的CytC含量明显增加,差异有统计学意义(P<0.01),见表1。
2.4 线粒体SOD活性和GSH、MDA含量测定 和常氧组相比,低氧组的线粒体SOD活性明显降低,GSH含量明显降低,MDA含量明显升高,差异有统计学意义(P<0.01),见表2。

表2 SOD活性和GSH、MDA含量
3 讨论
线粒体在细胞的正常生命活动和细胞死亡的过程中发挥着重要的作用,大多数的真核生物利用线粒体通过氧化磷酸化作用合成ATP,为机体各种生命活动提供能量。线粒体的正常功能与线粒体膜电位(mitochondrial transmembrane potential,Δ Ψm)有关,线粒体内外膜物质平衡依赖于线粒体的膜电位正常[6]。线粒体膜通透性(permeability transition,PT)改变可以引起ΔΨm的下降,而PT由MPTP所调控。MPTP是一种跨膜蛋白孔道,是由位于线粒体内膜和外膜上的多种蛋白组成的蛋白复合体。正常生理条件下,MPTP处于关闭状态,线粒体内膜除对一些选择性的代谢底物和离子外,对其他物质都没有通透性。然而在低氧、缺血再灌注损伤、钙超载等应激条件下,MPTP开放,通透性增加,内膜外的小分子物质大量进入内膜,使基质内的渗透压增大,又进一步促进小分子物质不断进入内膜,导致线粒体的肿胀,外膜破裂,从而释放存在于内外膜间的CytC和细胞凋亡诱导因子(apoptosis inducing factor,AIF)等凋亡蛋白,导致线粒体损伤和功能障碍[7-9]。Ca2+、氧化应激、无极磷酸盐、pH值增大等可以促进MPTP的开放,而Mg2+、环孢素A(CsA)、pH值降低、米帕林、狄布卡因可以抑制MPTP的开放[10]。MPTP开放的结果取决于开放的程度和时间,严重者可导致细胞凋亡或坏死[11]。
我们的先期实验发现慢性低氧高二氧化碳血症可以导致骨骼肌纤维萎缩,肌细胞线粒体增多并伴有线粒体内部结构的明显损害[12],凋亡可能部分参与了慢性低氧高二氧化碳引起的这种骨骼肌萎缩[13]。同时,我们还发现慢性低氧高二氧化碳条件下小鼠大脑线粒体能量生成障碍、COXI和III活性受到抑制,SOD活性、GSH含量显著降低,推测线粒体的功能障碍可能和氧化应激有关[14]。本实验采用离体的线粒体低氧模型,可以直接定位观察低氧对线粒体功能的损害。实验结果为低氧组MPTP的开放增加,线粒体膜电位下降明显,CytC释放增多,SOD活性、GSH含量显著降低,MDA含量明显升高。以上结果在某种程度上支持我们先期的实验结论。
线粒体是产生内源性自由基的主要场所,同时也是自由基攻击的靶部位[15]。本实验中MPTP开放和膜电位的下降,可能与自由基引起的损伤有关。与常氧组相比,低氧组小鼠骨骼肌离体线粒体中SOD活性、GSH含量显著降低,MDA含量明显升高。SOD和GSH是机体两种主要的自由基清除酶,其活性可以反映机体清除氧自由基的能力。GSH还是重要的保护因子,可以保护生物膜和生物大分子免受自由基的攻击。
氧自由基清除不足可以抑制SOD的活性,同时氧自由基还可以作用于生物膜中的多不饱和脂肪酸(polyunsaturated fatty acid,PUFA)引起脂质过氧化,并形成脂质过氧化物如MDA。线粒体产生ROS越多,对膜脂质攻击作用越强,生成的MDA也越多。MDA含量可以反映脂质过氧化的程度,间接反映了细胞受自由基攻击的严重程度。在本实验中,低氧组的小鼠骨骼肌离体线粒体内MDA含量明显升高,表明低氧后线粒体内氧自由基生成增加和组织损伤。因此,SOD活性、GSH含量下降,MDA含量增加,与氧化应激有关。这可能是MPTP开放增加,线粒体膜电位下降, CytC释放增加的重要原因。
此外,从理论上推测,MPTP开放,线粒体膜电位降低,CytC释放增加等还可诱导凋亡,从细胞凋亡途径影响线粒体。
综上所述,本实验从线粒体损伤角度解释了低氧所致机体损害的可能病理生理机制。低氧可通过增加小鼠肌肉离体线粒体的MPTP开放,下调膜电位,释放CytC,抑制SOD活性、降低GSH含量,升高MDA含量,即通过增强脂质过氧化反应和细胞凋亡途径等多种机制导致线粒体损伤。
[1] Peng YJ, Prabhakar NR. Effect of two paradigms of chronic intermittent hypoxia on carotid body sensory activity[J]. J Appl Physiol,2004,96(3):1236-1242.
[2] Katayama K, Sato Y, Morotome Y,et al. Intermittent hypoxia increases ventilation and SaO2during hypoxic exercise and hypoxic chemosensitivity [J]. J Appl Physiol, 2001,90(4):1431 -1440.
[3] Zoll J, Ponsot E,Dufour S,et al. Exercise training in normobaric hypoxia in endurance runners.III. Muscular adjustments of selected gene transcripts[J]. J Appl Physiol,2006,100 (4):1258-1266.
[4] Minezaki KK,Suleiman MS, Chapman RA. Changes in mitochondrial function induced in isolated guinea-pig ventricular myocytes by calcium overload[J].J Physiol,1994,476(3):459-471.
[5] Martinou JC,Desagher S,Antonsson B.Cytochrome C release from mitochondria :all or nothing[J].Nat Cell Biol,2000,2(3):41-43.
[6] Kroemer G,Zamzami N,Santos A.Mitochondrial control of apoptosis[J].Immunology Today,1997,18(1):44-51.
[7] Crompton M.The mitochondrial permeability transition pore and its role in cell death [J].Biochem J,1999,341(2):233-249.
[8] Halestrap AP.The mitochondrial permeability transition:its molecular mechanism and role in reperfusion injury[J].Biochem Soc Symp,1999,66:181-203.
[9] Halestrap AP,McStay GP,Clarke SJ.The permeability transition pore complex:another view [J].Biochimie,2002,84(2-3):153-166.
[10]Kim JS,He L,Lemasters JJ.Mitochondrial permeability transition:a common pathway to necrosis and apoptosis[J].Biochem Biophys Res Commun,2003,304(3):463-470.
[11]Chen M,Uerrero GAD,Huang L,et al.Caspase-9-induced mitochondrial disruption through cleavage of anti-apoptotic BCL-2 family members[J]. J Biol Chem,2007,282(46):33888-33895.
[12] 包绍智,房春燕,王小同,等.慢性低氧高二氧化碳对小鼠骨骼肌形态的影响[J].温州医学院学报,2009,39(1):8-11.
[13] 戴震宇,包绍智,黄汉津,等.凋亡在慢性低氧高二氧化碳小鼠骨骼肌萎缩中的作用[J].中国临床神经科学,2009,17(4):361-364.
[14] 吴彬,金露,王小同,等.慢性低氧高二氧化碳对小鼠大脑线粒体功能的影响[J].温州医学院学报,2010,40(2):115-118.
[15]Leeuwenburgh C,Heineeke JW.Oxidative stress and antioxidants in exercise [J].Curr Med Chem,2001,8(7):829-838.
猜你喜欢
世界科学技术-中医药现代化(2021年7期)2021-11-04
教育教学论坛(2019年19期)2019-06-17
山东林业科技(2019年2期)2019-06-03
天然产物研究与开发(2018年2期)2018-04-04
现代园艺(2017年11期)2017-06-28
奥秘(2016年10期)2016-12-17
中国运动医学杂志(2016年3期)2016-07-10
中国运动医学杂志(2016年3期)2016-07-10
中国卫生标准管理(2015年6期)2016-01-14
医学研究杂志(2015年5期)2015-06-10
