
家系、种子产生部位和土层深度对黄帚橐吾种子萌发出苗的影响
2010-12-24 00:52王满堂
草业科学 2010年8期
王满堂
(枣庄学院旅游与资源环境系,山东 枣庄277160)
近20多年来,青藏高原东北部高寒草甸退化严重,其显著特征是黄帚橐吾(Ligularia virgaurea)等毒杂草成份显著增加。调查显示,20世纪70年代末,黄帚橐吾盖度为5%~20%,而1992-1996年上升到40%~60%,草地品质下降[1]。单保庆等[1]、李小伟等[2]、刘左军等[3]对6个不同海拔样点的黄帚橐吾调查显示:均有大面积的斑块或与牧草混生群落,在斑块中黄帚橐吾均为绝对优势种,密度为60~120株/hm2,而在黄帚橐吾与牧草混生的群落中,密度为1~20株/hm2。由于对其生活史对策了解甚少,人们尚未找到治理黄帚橐吾的有效方法。有关种子萌发、出苗过程的研究成为生活史对策研究的一个重要内容,已受到生态学家的极大关注[4-5]。因为研究植物种子的萌发能力和萌发过程对环境条件的反应有助于理解植被组分种群动态的分布。种子的萌发、出苗受到诸多外因和内因的影响。环境因子影响种子萌发、出苗,比如土层深度通过调节土壤湿度、土壤温度和光照而调节种子萌发、出苗。种子大小的变异不仅存在于同一生境中物种间和同一物种不同种群间,还存在于同一种群不同个体间和同一个体不同部位间的层次。种子大小对种子萌发也能产生较大影响,Mckersie等[6]的研究结果表明,不同家系或者不同大小的黄帚橐吾种子,萌发能力有显著的不同;但是,在20℃下的萌发百分率与种子大小没有表现出一致的关系,同时种子大小对种子萌发速率没有影响,但是家系对萌发速率影响显著。
本试验主要研究了无光照条件下温度对黄帚橐吾种子萌发的影响,以及家系、种子产生部位和土层深度对黄帚橐吾种子出苗的影响,拟通过研究黄帚橐吾萌发、出苗过程,试图对其生态防除提供一些理论参考。
1 试验材料与方法
1.1 试验材料 黄帚橐吾隶属菊科橐吾属。黄帚橐吾为多年生草本,灰绿色,根肉质,簇生。在黄帚橐吾营养生长期,地上部分无茎,仅有1~10片簇生叶。黄帚橐吾既能进行有性繁殖,又能进行无性繁殖,是典型的游击型(guerilla)克隆生长植物。
1.2 温度对种子萌发影响 2004年9月中旬-10月上旬,在甘南州玛曲县城郊(35°58′N,101°53′E,海拔 3 500 m)随机采集黄帚橐吾种子。为保证所采种子已经成熟,在种子开始扩散时采集。去除败育、虫害的种子,并把所有种子混合在一起,装入牛皮纸信封。
温度模拟青藏高原东北部春季和夏季的气温,采用无光照变温,分3个处理:5/15,10/20,18/28 ℃,每天 12 h低温(5、10、18 ℃),12 h高温(15、20、28℃)。把种子置于直径为 9 cm的玻璃培养皿中进行培养,每套培养皿50粒种子,设置3个重复。培养皿底部垫有2层滤纸,以保证有充足的水分供应,每天加一次蒸馏水。种子萌发试验全部在E5型(加拿大产)植物生长培养箱中进行。
每天统计萌发的种子数,并把萌发的种子移走,同时每天加入适量的蒸馏水以保证滤纸湿润。萌发试验持续40 d,直到连续5 d不出现发芽的种子时终止统计。
1.3 土层深度对出苗的影响 于2005年4-10月兰州大学高寒草甸与湿地生态系统定位研究站(位于甘肃省合作市)进行土层深度对出苗影响的试验。合作市平均海拔为2 900 m,年平均气温 2.0℃,最冷的12、1、2三个月平均气温-8.9℃,最热的6-8月平均气温为11.5℃;年均降水量660 mm;属亚高山草甸植被类型[7]。
土层深度分 4个处理:0、1、3、5 cm。把 50粒种子播撒在花盆(高25 cm、直径30 cm)内,花盆内填充充分混匀的土壤(土壤早在2004年取自其自然生境,而且经薄膜覆盖数月消除土壤中的种子)。每个处理4个重复,共16盆。每天统计出苗数。
1.4 家系和种子产生部位对出苗的影响2004年 9-10月,在甘南州玛曲县城郊(35°58′N,101°53′E)、哇乐卡的不同种群中随机选定11株黄帚橐吾收集其种子(每株代表1个家系),每个种群只选1株进行有性繁殖的黄帚橐吾分株,种群之间相距至少5 m,尽量保证家系间的遗传差异。种子产生部位根据果实序的长度平均分为上、中、下3部分。试验于2005年4月开始,2005年10月结束。
2005年4月,为了研究家系、种子产生部位对种子大小的影响,将11个家系不同部位的种子用万分之一电子天平称量,然后把种子分别播撒在花盆内,花盆内填充充分混匀的土壤(土壤早在2004年取自其自然生境,并经薄膜覆盖数月消除土壤中的种子),种子上撒一层薄沙,有利保水,每盆播撒10粒种子,每个处理3个重复。每天统计出苗数。
1.5 数据处理

为了检验温度对萌发率的影响,使用一般线性模型(General Linear Model,GLM)中的ANOVA(方差分析)(温度为固定因素)进行分析,各数值经转化满足方差分析的要求。
出苗率=苗数/播种种子数×100%
采用GLM中的ANOVA(土层深度为固定因素)分析土层深度对出苗率的影响。采用GLM中的ANOVA(家系为随机因素)分析家系对种子大小、出苗率的影响,使用Mixed Model,把家系嵌套在种子产生部位中,即种子产生部位(家系)作为固定因素,进行ANOVA分析种子产生位置对种子大小、出苗率的影响。
2 结果与分析
2.1 不同温度下黄帚橐吾种子萌发 单因子方差分析结果表明,无光照下温度对黄帚橐吾种子出苗率有显著的影响(F=5.277,P=0.013)。温度越低,萌发率越高。5/15℃下黄帚橐吾种子萌发率为95.33%,显著大于18/28℃下黄帚橐吾种子萌发率(87.33%)(图1)。
2.2 土层深度对黄帚橐吾出苗的影响 单因子方差分析结果表明,土层深度对黄帚橐吾种子出苗率有显著的影响(F=12.121,P=0.002)。土层越深,出苗率越低。土层深度为0 cm时,出苗率最高,达61.66%;土层深度为1 cm时,出苗率为42.50%;土层深度为3 cm时,出苗率锐减,为0.83%;土层深度5 cm时,黄帚橐吾出苗率为0(图2)。
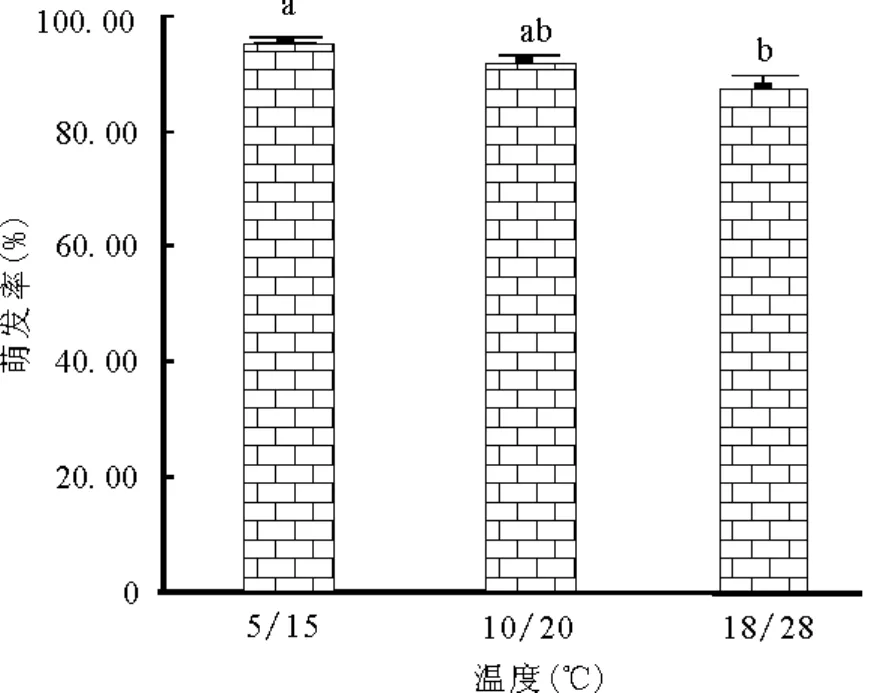
图1 不同温度下黄帚橐吾种子萌发率
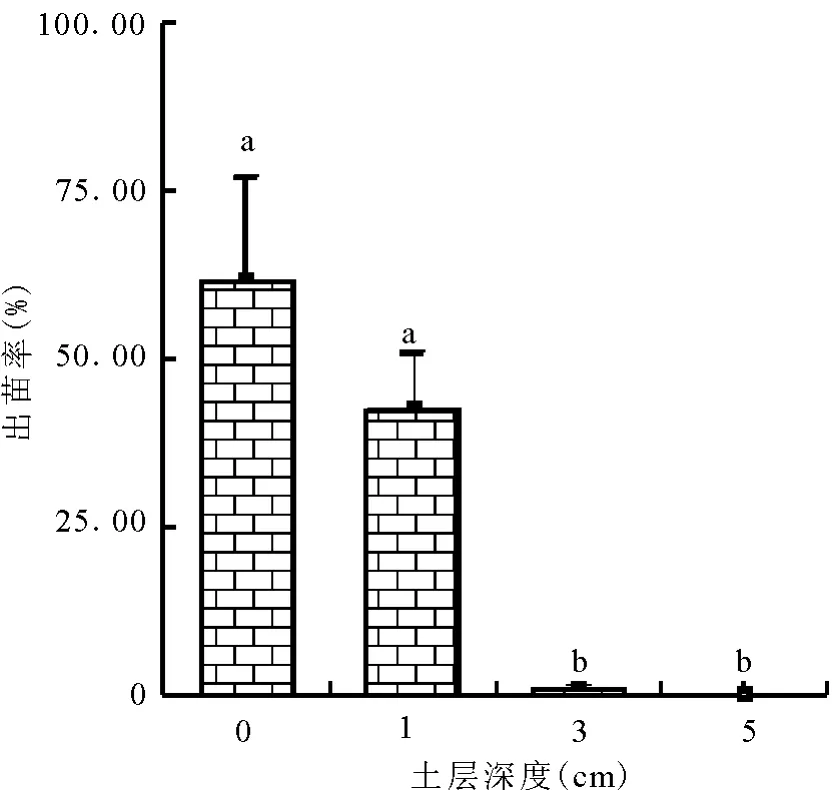
图2 不同土层深度黄帚橐吾种子出苗率
2.3 家系和种子产生部位对黄帚橐吾种子大小和出苗率的影响 研究表明,家系对黄帚橐吾种子大小、种子出苗率有显著的影响,说明种子萌发能力有遗传基础。由于种子大小、出苗率受到家系的影响,因而,在分析种子产生部位对种子大小和出苗率的影响时,使用混合模型,构建了种子产生部位(家系),用同一个家系不同种子产生部位进行比较。结果表明,种子产生部位(家系)对种子大小、出苗率也有显著影响(图 3,表1)。不同家系种子大小不同,出苗率相差很大,大种子出苗率高。同一家系不同种子产生部位种子大小不同,出苗率也不同。黄帚橐吾顶部种子最先成熟,底部的种子最后成熟,对种子大小和出苗率产生影响。
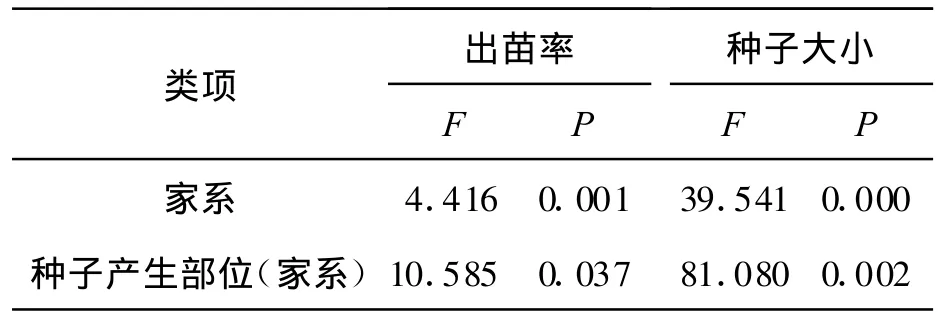
表1 不同家系、种子产生部位对黄帚橐吾种子大小和出苗率的方差分析
3 讨论
结合黄帚橐吾种子其他室内萌发试验结果[8-9],可以发现黄帚橐吾种子没有休眠,在光照和黑暗条件下,种子均可以萌发,黄帚橐吾种子萌发率是大于80%。然而由土层深度对黄帚橐吾种子出苗影响的试验结果和其他盆栽试验[9]结果可以看出,黄帚橐吾种子出苗率明显小于室内种子萌发率。这可能是由于病毒、细菌的感染导致种子或胚根腐败,或者种子被一些土壤中的昆虫吃掉[10-11]。
与其他植物一样[12],黄帚橐吾种子出苗率受到土层深度的影响,随着土层深度增加出苗率降低。黄帚橐吾种子在其生境的温度范围内有很高的萌发率。在青藏高原高寒草甸,持久土壤种子库中没有发现黄帚橐吾,瞬时土壤种子库只有很少量的黄帚橐吾[13],说明黄帚橐吾种子对土壤种子库的贡献不大。黄帚橐吾种子在高寒草甸物种中属于小种子物种,小种子储存的物质不足以使种子从很深的土层破土。故可以推测随着土层深度增加到3 cm以上出苗率低,其主要原因是萌发的种子不能破土从而死掉。以上研究结果,是解释很难在野外观测到黄帚橐吾实生苗的原因之一。
通过研究发现,黄帚橐吾种子产生部位对黄帚橐吾种子大小和出苗率具有显著的影响。黄帚橐吾顶部种子最先成熟,底部的种子最后成熟,对种子出苗率产生影响。因此种子出苗能力不仅受到遗传结构控制,与种子成熟早晚也有关系。许多研究发现在生产多个花的物种中,随着花的开放次序(较早开放-较晚开放),植物对种子的投入会降低[14]。较早开放的花往往能抢先占有资源,这符合“资源竞争假说”[15]。黄帚橐吾总状花序、不同部位头状花序在资源投入上受其栖息生境各种生态因子的影响,总状花序内的资源分配存在着位置依赖性,表现为顶部头状花序比基部和中部头状花序有较大的资源分配[3]。黄帚橐吾果实成熟顺序是从顶部到底部,并且随着果实成熟,承载这部分果实的茎及上面的叶枯萎。因而由于种子从母体获取资源的不同,不同部位的种子大小有显著的差异。此外,生境和种群密度对不同家系间的种子大小有极显著的影响[16],表明黄帚橐吾种子大小变化有遗传基础。因此,黄帚橐吾种子大小不仅受个体发育的影响(即产生种子的时间差异),而且受遗传因素影响。家系对种子出苗率有显著的影响,说明种子萌发能力有遗传基础。黄帚橐吾种子产生部位对种子出苗率也有显著的影响。因此种子萌发、出苗能力不仅受到遗传结构控制,与种子成熟早晚也有关系。
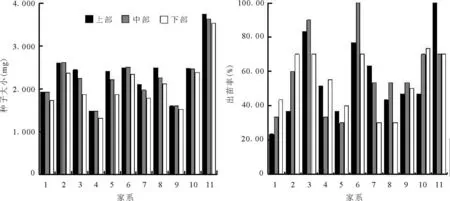
图3 不同家系、种子产生部位黄帚橐吾种子大小和出苗率
黄帚橐吾不仅能够由克隆生长产生分株。对于一个植物种群而言,有性繁殖比无性繁殖更重要,因为有性繁殖能够产生具有新的适应特征的基因型,从而增加植物的生存和竞争能力,并且能够长距离扩散从而增加其蔓延的速度和范围。研究发现,通过土层深度、家系和种子产生部位对黄帚橐吾种子萌发出苗的影响,可以发现大种子更容易萌发、出苗,而生境和种群密度对不同家系间黄帚橐吾种子大小产生影响,因而可以通过对生境和种群密度的干扰从而影响黄帚橐吾种子大小,达到控制黄帚橐吾的目的。
[1]单保庆,杜国祯,刘振恒.不同养分条件下和不同生境类型中根茎草本黄帚橐吾的克隆生长[J].植物生态学报,2000,24(1):46-51.
[2]李小伟,孙坤,马瑞君,等.甘南州天然草地有毒植物及其防治对策[J].草业科学,2003,20(10):60-63.
[3]刘左军,杜国祯,陈家宽.影响黄帚橐吾种子生产的因素Ⅰ:生境和花序结构[J].植物生态学报,2003,27:344-351.
[4]刘志民,蒋德明,高红瑛,等.植物生活史繁殖对策与干扰关系的研究[J].应用生态学报,2003,14:418-422.
[5]Bu H Y,Chen X L,Xu X L,etal.Seed size and germination in an alpine meadow on the eastern Qinghai-Tibet plateau[J].Plant Ecology,2007,191:127-149.
[6]Mckersie B D,Tomes D T,Yamamoto S.Effect of seed size on germination,seedling vigor,electrolyte,and establishment of bird's-foot trefoil[J].Canadian Journal of Plant Science,1981,61:337-343.
[7]吴征镒.中国植被[M].北京:科学出版社,1980.
[8]王满堂.青藏高原高寒草甸黄帚橐吾生活史对策研究[D].兰州:兰州大学,2007.
[9]Wang M T,Lu N N,Zhao Z G.Effects of temperature and storage length on seed germination and the effects of light conditions on seedling establishment with respect to seed size in Ligularia virgaurea[J].Plant Species Biology,2009,24:120-126.
[10]Chambers JC,MacMahon JA.A day in the life of a seed:Movements and fates of seeds and their implication for natural and managed systems[J].Annual Review of Ecology,Evolution,and Systematics,1994,25:263-292.
[11]Fenner M,Thompson K.The Ecology of Seeds[M].Cambirdge:Cambridge University Press,2005.
[12]Benvenuti S,Macchia M,Miele S.Light,temperature and burial depth effects on Rumex obtusif olius seed germination and emergence[J].Weed Research,2001,41:177-186.
[13]周显辉.青藏高原高寒草甸土壤种子库研究[D].兰州:兰州大学,2006.
[14]Luisa B M.Intra-inflorescence variation in floral traits and reproductive success of the hermaphrodite Silene acutifolia[J].Annals of Botany,2004,94:441-448.
[15]Thomson J D.Deployment of ovules and pollen among flowers within inflorescences[J].Evolutionary Trends in Plants,1989(3):65-68.
[16]刘左军,杜国祯,陈家宽.不同生境下黄帚橐吾个体大小依赖的繁殖分配[J].植物生态学报,2002,26:44-50.
猜你喜欢
辽宁农业科学(2021年5期)2021-11-03
落叶果树(2021年6期)2021-02-12
新农业(2018年6期)2018-04-18
现代农业(2016年5期)2016-02-28
体育科技(2016年2期)2016-02-28
广东海洋大学学报(2015年4期)2016-01-13
听力学及言语疾病杂志(2015年5期)2015-12-24
首都医科大学学报(2015年4期)2015-12-16
西华师范大学学报(自然科学版)(2015年3期)2015-02-27
郑州大学学报(医学版)(2015年2期)2015-02-27
