
贵州省PRRSV分离株Nsp2基因的克隆及序列分析
2010-09-11 07:21王开功周碧君罗险峰安同庆刘春国程振涛路宪礼
中国预防兽医学报 2010年12期
刘 飞,王开功,*,周碧君,,罗险峰,文 明,,安同庆,刘春国,程振涛,路宪礼,陈 俊
(1.贵州大学动物科学学院,贵州贵阳550025;2.贵州省动物疫病研究室,贵州贵阳550025;
3.贵州省动物疾控中心,贵州贵阳550008;4.中国农业科学院哈尔滨兽医研究所,黑龙江哈尔滨150001)
猪繁殖与呼吸综合征(Porcine reproductive and respiratory syndrome,PRRS)是由PRRS病毒(PRRSV)引起的一种高度传染性疾病。该病自1987年首次在美国被报道发生以来,迅速蔓延至各养猪国家,并造成了严重的经济损失,是目前危害养猪业的重要疾病之一[1-2]。1996年郭宝清等首次在国内分离到PRRSV,证实了该病在我国的存在[3]。PRRSV基因组长约15kb,有9个开放阅读框(ORF)[4-6]。其中ORF1a和ORF1b约占全基因组的80%,编码病毒RNA复制酶多聚蛋白,并在自剪切蛋白酶的作用下产生13个非结构蛋白(Nsp1α、N sp1β、N sp2~Nsp12),其中Nsp2蛋白变异性很大[7-8]。
2006 年以来我国暴发了高致病性PRRS,病原为高致病性PRRSV(HP-PRRSV),其主要分子特征为Nsp2蛋白存在30个氨基酸的不连续缺失,即第481位和533位~561位氨基酸缺失[2,9]。这一缺失特征普遍存在于2006年~2009年分离的HP-PRRSV中,并且高度保守[10]。迄今为止,仍无HP-PRRSV Nsp2蛋白发生进一步扩大缺失的报道。但是,通过对2007年~2009年贵州省不同地区的HP-PRRSV分离株的Nsp 2蛋白进行分析,我们发现部分HP-PRRSV的Nsp2蛋白缺失正在扩大。这些新缺失病毒株的出现,提示了HP-PRRSV可能出现了新的变异走向。
1 材料和方法
1.1 病毒样品和细菌株 2007年~2009年本实验室在贵州省的贵阳、花溪、遵义、毕节和铜仁等地区采集疑似PRRSV感染的临床样品12份,其中11份鉴定为阳性[11];大肠杆菌感受态细胞DH5α株由本实验室保存。
1.2 主要试剂 AMV RTase购自上海生工生物工程技术服务有限公司;RNAiso Reagent、Taq酶等PCR试剂、PstⅠ和EcoRⅠ内切酶、pMD18-T载体均购自TaKaRa公司;胶回收试剂盒购自Omega公司;质粒提取试剂盒购自博大泰恒公司。
1.3 PRRSV Nsp2基因的扩增 参考GenBank中登录的HP-PRRSV的基因序列,在Nsp2基因内部的保守区设计1对引物:PNsp2-1:5'-CCTCCGTGGTG CAACAAATCTTG-3'; PNsp2-2: 5'-CGATGATGGCT TGAGCTGAGTAT-3',扩增片段大小为1064bp。引物由上海生工生物工程技术服务有限公司合成。
按照常规方法提取病毒RNA并进行RT-PCR扩增。PCR反应体系为 50μL:cDNA 5μL、10×PCR Buffer 5μL、MgCl2(25mmol/L)3μL、dNTPs(10mmol/μL)1μL、Taq DNA 聚合酶(2.5U/μL)1μL、上下游引物(20μmol/μL)各 1μL和灭菌双蒸水33μL。扩增条件为:94℃ 5m in;94℃ 30s、56℃ 30s、72℃ 45s,35个循环;72℃ 10min。PCR产物进行琼脂糖凝胶电泳观察结果。
1.4 目的基因的克隆与鉴定 将目的基因片段胶回收后连接至pMD18-T载体中,并转化DH5α感受态细胞,涂布LB/Amp+/IPTG/X-gal平板,蓝白斑筛选后提取重组质粒DNA。经PCR、PstⅠ/EcoRⅠ双酶切鉴定,阳性重组质粒由TaKaRa公司测序。
1.5 Nsp2基因序列的比对分析 按照Cha等建立的方法进行Nsp2基因序列的分析[12]。采用LasergeneR7.0(DNAStar)和DNAMan软件,对贵州HP-PRRSV分离株Nsp2基因序列与GenBank登录的52株国内外代表性病毒株序列进行多重序列比对,采用MEGA4软件根据neighbor-joining方法绘制系统发生树。
2 结果
2.1 RT-PCR扩增结果 11份HP-PRRSV阳性病料样品的RNA经RT-PCR扩增后,凝胶电泳结果显示在约1kb处出现扩增条带,虽与预期的1064bp相近,但却存在大小不同的两条泳带。泳道10、11的片段比泳道1~8的稍小,在泳道9中同时存在这两种大小的片段,提示可能在同一份病料样品中同时存在两种不同缺失类型的PRRSV(图1)。
2.2 Nsp2基因片段的克隆与鉴定 将Nsp2部分基因片段的PCR产物连接至pMD18-T载体中,提取重组质粒进行PCR和PstⅠ/EcoRⅠ双酶切鉴定,选择阳性重组质粒进行测序。其中GZHW泳道的两条带分别切胶连接至pMD18-T载体中进行测序。结果显示GZHW的两条带分别为1064bp(GZHW 1)和977bp(GZHW 2),表明病猪混合感染了2种Nsp2基因不同程度缺失的PRRSV。另外的10份样品中有7份样品(GZCJ、GZHX、GZDJ、GZJS、GZWB、GZKY和GZKB)的Nsp2基因扩增片段大小为1064bp,1份样品(GZZB)扩增片段为1061bp,2个样品(GZWM、GZLJ1)的扩增片段为977bp。12株病毒的序列已录入GenBank(EU140613~EU140624)(表 1)。
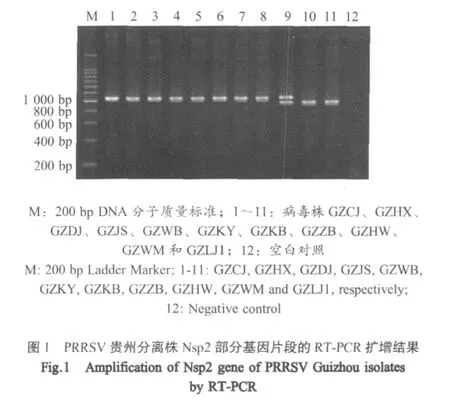
表1 HP-PRRSV贵州分离株信息Table 1The information of HP-PRRSV isolates of Guizhou area
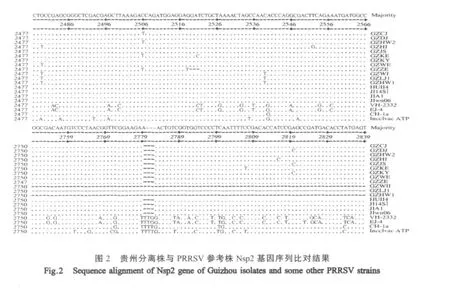
2.3 Nsp2基因序列分析 将12株分离株的Nsp2基因序列与GenBank中登录的参考毒株VR2332、BJ-4等的序列进行多重比对显示:GZCJ等8株HP-PRRSV发生2处不连续缺失,缺失位置为2780bp~2782bp和2936bp~3022bp,共 90个碱基,与以JXA1为代表的HP-PRRSV缺失位置相同。GZHW 2、GZWM和GZLJ1同样发生2处不连续缺失,但缺失位置为2750bp~2839bp和2936bp~3022bp,共177个碱基,是一种新型Nsp2基因缺失型PRRSV株。GZZB株则发生3处不连续缺失,缺失位置为2519bp~2521bp、2780bp~2782bp和2936bp~3022bp,共93个碱基,也是一种新型缺失病毒株(图2)。
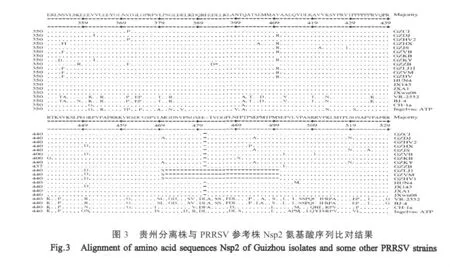
2.4 Nsp2氨基酸序列分析 将根据12株HPPRRSV贵州分离株Nsp2基因推导的氨基酸序列与VR2332、BJ-4株等比较显示:GZCJ等8株分离株Nsp2蛋白在第481位和533位~561位发生缺失,共缺失30个氨基酸,与2006年江西省分离的PRRSV JXA1株的Nsp2氨基酸缺失情况完全相同(图 3);GZHW 2、GZWM 与 GZLJ1分离株 Nsp2蛋白缺失471位~500位和533位~561位,共59个氨基酸;GZZB分离株Nsp2基因缺失第394位、第481位和533位~561位,共31个氨基酸。

2.5 Nsp2序列的系统发生树分析 将PRRSV贵州分离株与其他地区PRRSV分离株的Nsp2氨基酸序列进行系统发生树分析。结果表明:贵州分离株与其他HP-PRRSV变异株(如JXA1、HuN4等)位于同一大的分支中,亲缘关系较近,而与国内2006年以前分离的经典病毒株的亲缘关系相对较远;同时,贵州分离株可分为两个小的分支,分支1与CBB-1-F3同源性最高,分支2与SX2009同源性最高(图4)。同源性分析结果显示:12株贵州分离株之间Nsp2核苷酸序列同源性为97.2%~100%,氨基酸同源性为94.9%~100%;与国内近来流行的变异株的核苷酸同源性为96.8%~99.3%,氨基酸同源性为93.8%~98.9%;而与早期国内外分离的经典病毒株的核苷酸同源性为75.1%~98.5%,氨基酸同源性为65.8%~97.7%。
3 讨论
通过对贵州省PRRSV分离株的序列分析表明,Nsp2基因不同程度的缺失是贵州省PRRSV变异的明显特征。在测定序列的12株分离株中,同一份病料样品中存在2种Nsp2基因不同程度缺失的PRRSV(GZHW 1和GZHW 2),提示 PRRSV有进一步扩大缺失的趋势。
本研究12株分离株中共有3种不同缺失型:以GZKB株为代表的Nsp2蛋白30个氨基酸缺失型,即在氨基酸序列第481位和第533位~第561位发生2处共30个氨基酸不连续缺失。该型分离株为贵州省的主要流行类型,与2006年引起我国HP-PRRS的病毒株及国内其它地区分离株缺失位点完全一致[2]。该缺失特征为HP-PRRSV的重要标志,是判断PRRSV是否为高致病性病毒株的重要指标[13],而本研究中12株PRRSV贵州分离株均为HPPRRSV。
以GZZB株为代表的Nsp2蛋白31个氨基酸缺失型,在30个氨基酸缺失型基础上,在第395位又缺失一个氨基酸。31个氨基酸缺失型为一种新型PRRSV Nsp2缺失型病毒株,但所占比例较小,不是贵州省的主要流行株类型。
以GZWM株为代表的Nsp2蛋白59个氨基酸缺失型,既在30个氨基酸缺失型基础上,在Nsp2蛋白的第481位氨基酸的两翼又缺失了29个氨基酸,是一种新的HP-PRRSV突变株。本研究中共分离3株该类型PRRSV,并且仅在贵州省分离获得。
研究表明,Nsp2蛋白是PRRSV非结构蛋白区域变异最大的,并且变异株间Nsp2保守性较差,抗原表位较多,而表位集中的区域容易发生缺失或突变[14]。由于PRRSV不断发生变异,对其基因序列进行遗传变异分析有助于设计和开发更有效的PRRSV诊断制剂,早期预警PRRS的发生和流行。本研究获得的新缺失型分离株,是否为适应性变异,能否承受内外环境的选择压力,有待于进一步调查研究。
[1]Colines JE,Benfield D A,Christianson W T,et al.Isolation of swine infertility and respiratory syndrome virus(isolate ATCC VR-2332)in North America and experimental reproduction of the disease in gnotobiotic pigs[J].JVet Diagn Invest,1992,4:117-126.
[2]童光志,周艳君,郝晓芳,等.致病性猪繁殖与呼吸综合征病毒的分离鉴定及其分子流行病学分析[J].中国预防兽医学报,2007,29(5):323-327.
[3]郭宝清,陈章水,刘文兴,等.从疑似PRRS流产胎儿分离猪生殖和呼吸综合征病毒(PRRSV)的研究[J].中国畜禽传染病,1996,18(2):1-5.
[4]Conzelmann K K,Visser N,Van Woensel P,et al.Molecular characterization of porcine reproductive and respiratory syndrome virus,a member of the arterivirus group[J].Virology,1993,193(1):329-339.
[5]Meulenberg J J,Hul St M M,De Meijer E J,et al.Lelystad virus,the causative agent of porcine epidemic abortion and respiratory syndrome(PEARS),is related to LDV and EAV[J].Virology,1993,192(1):62-72.
[6]Murtaugh M P,Elam M R,Kakach L T.Comparison of the structural protein coding sequences of the VR-2332and Lelystad virus stains of the PRRS virus[J].Arch Virol,1995,140:1451-1460.
[7]Gao Zhi-Qiang,Guo Xin,Yang Han-Chun.Genom ic characterization of two Chinese isolates of porcine respiratory and reproductive syndrome virus[J].Arch Virol,2004,149:1341-1351.
[8]Yoshii M,Okinaga T,M iyazaki A,et al.Genetic polymorphism of the nsp2gene in North American type porcine reproductive and respiratory syndrome virus[J].Arch Virol,2008,153(7):1323-1334.
[9]Tian K,Yu X,Zhao T,et al.Emergence of fatal PRRSV variants:unparalleled outbreaks of atypical PRRS in China and molecular dissection of the unique hallmark[J].PLoS ONE,2007,6(13):526.
[10]Zhou L,Chen S,Zhang J,et al.Molecular variation analysis of porcine reproductive and respiratory syndrome virus in China[J].Virus Res,2009,145(1):97-105.
[11]毛君婷,周碧君,史开志,等.PRRSV不同NSP2基因缺失型毒株混合感染的鉴定[J].中国兽医科学,2009,39(02):150-153.
[12]Cha S H,Choi E J,Park J H,et al.Molecular characterization of recent Korean porcine reproductive and respiratory syndrome(PRRS)viruses and comparison to other Asian PRRS viruses[J].Vet M icrobiol,2006,117:248-257.
[13]Zhou L,Zhong J L,Zeng JW,et al.The 30-Am ino-Acid deletion in the Nsp2of highly pathogenic porcine reproductive and respiratory syndrome virus emerging in China is not related to its virulence[J].JVirol,2009,83(10):5156-5167.
[14]万根,许兰娇,邓舜洲,等.高致病性PRRSV NSP2基因的扩增与分析[J].江西农业大学学报,2009,31(4):737.
猜你喜欢
贵州畜牧兽医(2022年6期)2022-12-29
趣味(数学)(2020年4期)2020-07-27
支部建设(2020年15期)2020-07-08
中国洗涤用品工业(2019年4期)2019-05-11
自然资源情报(2018年8期)2018-12-28
中成药(2018年1期)2018-02-02
领导决策信息(2017年17期)2017-06-21
中成药(2017年3期)2017-05-17
百科知识(2015年18期)2015-09-10
中国交通信息化(2015年11期)2015-06-06
