
H9N2亚型禽流感病毒的遗传进化分析及F株灭活疫苗的免疫效果评价
2010-09-11 07:21童海兵张小荣吴艳涛刘学贤龚建森刘秀梵
中国预防兽医学报 2010年12期
童海兵,张小荣,吴艳涛,刘学贤,龚建森,刘秀梵*
(1.扬州大学兽医学院,江苏扬州225009;2.中国农业科学院家禽研究所,江苏扬州225009)
禽流感是由A型流感病毒引起的禽类急性呼吸道传染病。禽流感病毒(Avian influenza virus,AIV)属于正粘病毒科,是一种分8个节段的单股、负链RNA病毒。病毒粒子囊膜表面分布血凝素(HA)和神经氨酸酶(NA)两种糖蛋白,目前已经发现16种HA亚型和9种NA亚型[1]。H9N2亚型AIV对于禽类为低致病性,但在混合感染或继发感染其他病原的情况下,仍可引起高发病率和死亡率[2-3]。由于我国普遍使用H9亚型禽流感灭活疫苗,免疫压力下病毒变异加快,出现多个基因亚型的H9N2亚型AIV流行毒株[4-6]。本研究对2009年~2010年期间从免疫鸡群中分离的H9N2亚型AIV毒株进行HA基因序列测定和分析,以了解病毒的变异情况,并评价了用于制备H9亚型F株禽流感灭活疫苗对分离毒株的免疫效果。
1 材料和方法
1.1 H9N2亚型AIV毒株 9个分离毒株为:A/Chicken/Anhui/L9/2009(简称 L9株)、A/Chicken/Fujian/F9/2009(简称 F9株)、A/Chicken/Fujian/F119/2009(简称 F119株)、A/Chicken/Fujian/S9/2009(简称S9株)、A/Chicken/Henan/YD/2009(简称 YD 株)、A/Chicken/Jilin/D9/2009(简称 D9株)、A/Chicken/Zhejiang/M/2009(简称 M 株)、A/Chicken/Anhui/W 9/2009(简称 W 9株)、A/Chicken/Jilin/D119/2009(简称 D119株),这些毒株是于2009年~2010年期间从接种过H9亚型F株禽流感灭活疫苗的鸡群中分离。疫苗毒株为A/chicken/Shanghai/F/98(F株)[6],由扬州大学农业部畜禽传染病学重点开放实验室提供。
病毒半数鸡胚感染量(EID50)的测定方法为:将病毒用灭菌PBS作10倍稀释,取10-5~10-106个稀释度,尿囊腔接种5个10日龄SPF鸡胚(购自北京梅里亚维通公司),37℃培养72h后,测定每个鸡胚尿囊液HA价,利用Reed-Muench法计算EID50。
1.2 主要试剂 TIANamp病毒RNA提取试剂盒购自天根生化科技(北京)有限公司;AxyPrep PCR清洁试剂盒和AxyPrep DNA凝胶回收试剂盒购自爱思进生物技术(杭州)有限公司;Expand High Fidelity PCR System、dNTPs(10mmol/μL)等购自 Roche 公司;T4DNA连接酶、AMV反转录酶(10u/μL)、RNasin(40u/μL)购自Promega公司;pCR2.1载体购自Invitrogen公司;H9亚型F株禽流感灭活疫苗购自扬州威克生物工程有限公司(生产批号:1002003)。
1.3 引物设计和合成 参照文献[7]的方法设计反转录(RT)引物Uni12-primer(5'-AGCAAAAGCAG G-3'),参照GenBank中登录的AIV序列设计PCR引物H9HA-F(5'-AGCAAAAGCAGGGGWANTTCAC AA-3')和H9HA-R(5'-GCCAATTATATACAAATGTT GCATCTGC-3')。所有引物均由南京金斯特生物科技有限公司合成。
1.4 RT-PCR扩增HA基因 取140μL含病毒鸡胚尿囊液,用试剂盒提取病毒RNA。用Uni12-primer引物和AMV反转录酶进行cDNA合成,反应体系为30μL,RNA用量为500ng,42℃反应60m in。
取2μL cDNA进行PCR,反应体系为:10×Expand High Fidelity Buffer w ith 15mM MgCl25μL、dNTPs(10mmol/μL)1μL、Expand High Fidelity Enzyme M ix 1μL、RNasin 1μL、引物 H9HA-F和H9HA-R各25pmol,用纯水补足至总体积50μL。反应条件:94℃5min;94℃30s、60℃30s、72℃1.5m in;共30个循环,72℃8m in。
PCR产物用AxyPrep PCR清洁试剂盒和AxyPrep DNA凝胶回收试剂盒进行纯化,再以T/A克隆的方法插于pCR2.1载体中。
1.5 HA基因的测序和遗传进化树绘制 含HA基因的重组质粒由南京金斯特生物科技有限公司测序。采用Lasergene 7.1软件进行HA基因序列编辑和翻译;用Mega4.0.2软件绘制遗传进化树。
1.6 F株灭活疫苗对分离株的免疫保护试验 将200只3周龄SPF鸡(种蛋购自北京梅里亚维通公司,自行孵化)随机分为20组,每组10只,隔离饲养;其中10组接种H9亚型F株禽流感灭活疫苗(0.3m L/只),另外10组接种灭菌PBS(0.3m L/只)作为对照。21d后采集各组鸡的血清,测定血凝抑制(HI)抗体效价(以log2表示);分别用9个分离毒株和F株经鼻腔途径攻毒,每个病毒株攻击疫苗接种组和对照组各1组,剂量为每只鸡106EID50/0.1m L。攻毒后第5d,采集喉头和泄殖腔拭子样品,每只鸡的样品接种3枚10日龄SPF鸡胚,37℃培养72h后测定每个鸡胚尿囊液HA价。分别统计各组鸡的排毒情况。
2 结 果
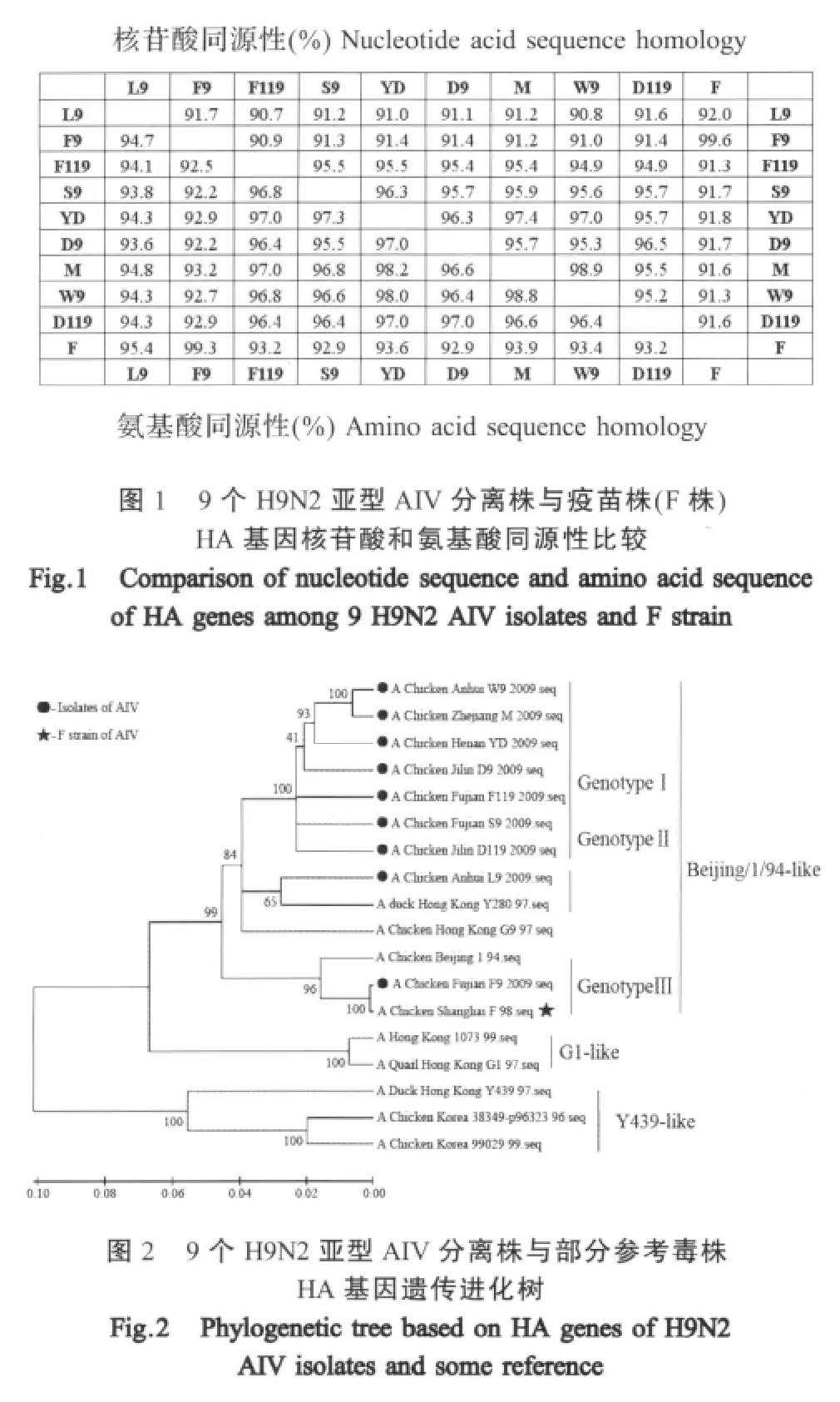
2.1 HA基因的序列分析 分别对9个分离株HA基因测序结果显示其编码区长度均为1683nt;同源性分析表明;9个分离株之间核苷酸序列同源性为90.7%~98.9%,推导氨基酸序列同源性为92.2%~98.8%;分离株与F株之间HA的核苷酸序列同源性为91.6%~99.6%,氨基酸序列同源性为92.9%~99.3%(图1)。遗传进化分析证实,9个分离毒株与F株均属于Beijing/1/94-like分支,但分属3个不同的基因亚型(图2)。
2.2 F株的免疫效果评价 3周龄SPF鸡接种H9亚型F株禽流感灭活疫苗21d后,产生的HI抗体效价在9Log2以上,对分离株及F株攻毒后的喉头和泄殖腔排毒产生明显的抑制作用(表1)。其中L9株、F9株、S9株、D9株、M株、D119株和F株攻毒后5d均未检出排毒,F119株、YD株、W 9株攻毒后5d的排毒率仅为1/10,而未接种疫苗的对照组攻毒后5d排毒率均为10/10。
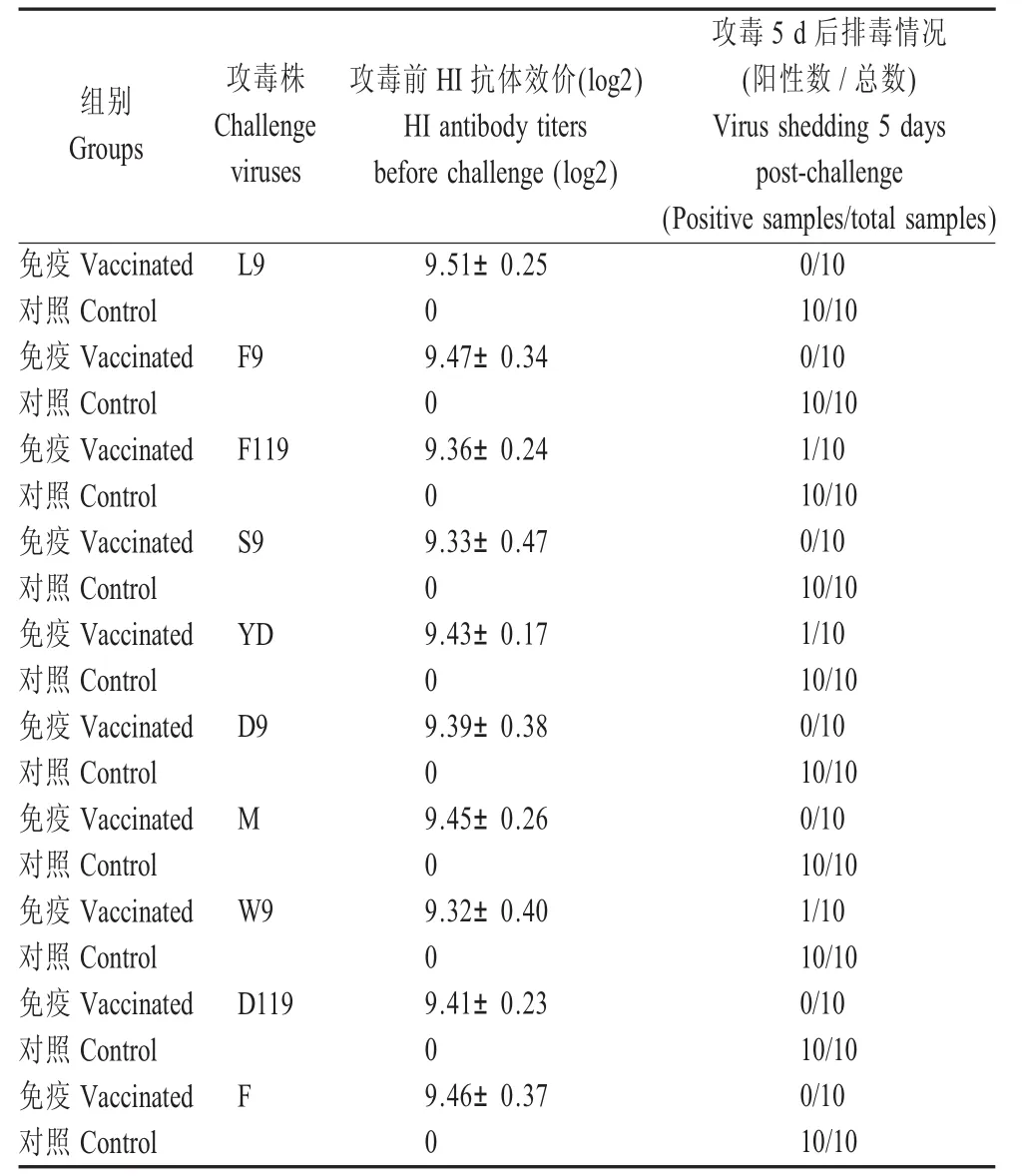
表1 H9亚型禽流感灭活疫苗(F株)对9个分离株的免疫保护效果Table 1Protection efficacy of H9subtype F strain avian influenza inactivated vaccine against 9isolates
3 讨论
自上世纪90年代后期以来,H9N2亚型AIV在许多国家的家禽(特别是鸡群)中广泛流行。陆生家禽中分离的H9N2亚型AIV毒株有3个进化分支[4,8-14]:G1分支主要在鹌鹑中被发现(代表毒株是A/Quail/HongKong/G1/1997);Y280分支在多种禽类中均有流行(代表毒株是A/Duck/HongKong/Y280/1997),该分支也被称为Beijing/1/94-like(代表毒株是A/Chicken/Beijing/1/94);Y439分支的代表毒株是 A/Duck/Hong Kong/Y439/97。3个分支的病毒在欧亚大陆家禽中流行,并进化为多个基因亚型。防控H9N2亚型AIV感染的重要措施是采用同亚型的疫苗进行免疫接种,但由于流感病毒易发生变异,必须掌握流行病毒株的变异情况。近年来,国内有报道认为当前流行病毒株已经发生了显著变化,现有疫苗已经不能产生完全免疫保护[15]。也有研究认为虽然H9亚型AIV发生了一定变异,但是现有疫苗仍能提供有效保护[16]。根据本研究对2009年~2010年期间从H9亚型F株禽流感灭活疫苗免疫鸡群分离的流行毒株所进行的HA基因测序分析,分离株HA基因核苷酸序列同源性为90.7%~98.9%,氨基酸序列同源性为92.2%~98.8%;分离株与F株HA之间的核苷酸序列同源性为91.6%~99.6%,氨基酸序列同源性为92.9%~99.3%。9个分离株又可以分为3个基因亚型,其中F9分离株与F株属于同一基因亚型,L9分离株与A/Chicken/Hong Kong/G9/97株属于同一基因亚型,另外7个分离株属于独立的一个基因亚型,表明当前H9亚型AIV流行毒株出现较大变异。但是,9个分离毒株与F株均属于Beijing/1/94-like进化分支,未发现G1-like和Y439-like分支,表明当前的流行毒株具有同一起源[4,6],这对应用疫苗预防H9亚型AIV感染是有利的。
由F株制备的H9亚型禽流感灭活疫苗在国内的应用范围非常广泛。本研究发现F株对9个分离株均能提供很高的保护率,表明当前的流行毒株虽然在基因水平上发生了变异,但是并未造成太大的抗原性的变化,该结果与荣骏弓等和姜北宇等的结果一致[16-17]。尽管如此,由于流感病毒本身的易变异性,再加上疫苗广泛使用造成的免疫选择压力,H9亚型AIV的变异仍将继续,因此,今后仍需继续监测病毒的变异情况,为筛选新的疫苗毒株作好准备。
[1]Fouchier R A,Munster V,Wallensten A,et al.Characterization of a novel influenza A virus hemagglutinin subtype(H16)obtained from black-headed gulls[J].JVirol,2005,79(5):2814-2822.
[2]Haghighat-Jahrom i M,Asasi K,Nili H,et al.Coinfection of avian influenza virus(H9N2subtype)w ith infectious bronchitis live vaccine[J].Arch Virol,2008,153(4):651-655.
[3]Kishida N,Sakoda Y,Eto M,et al.Co-infection of Staphylococcus aureus or Haemophilus paragallinarum exacerbates H9N2influenza A virus infection in chickens[J].Arch Virol,2004,149(11):2095-2104.
[4]Li Chengjun,Yu Kangzhen,Tian Guobin,et al.Evolution of H9N2influenza viruses from domestic poultry in Mainland China[J].Virology,2005,340(1):70-83.
[5]Choi Y K,Ozaki H,Webby R J,et al.Continuing evolution of H9N2influenza viruses in Southeastern China[J].J Virol,2004,78(16):8609-8614.
[6]Liu Hongqi,Liu Xiufan,Cheng Jian,et al.Phylogenetic analysis of the hemagglutinin genes of twenty-six avian influenza viruses of subtype H9N2isolated from chickens in China during 1996-2001[J].Avian Dis,2003,47(1):116-127.
[7]Hoffmann E,Stech J,Guan Y,et al.Universal primer set for the full-length amplification of all influenza A viruses[J].Arch Virol,2001,146(12):2275-2289.
[8]Homayounimehr A R,Dadras H,Shoushtari A,et al.Sequence and phylogenetic analysis of the haemagglutinin genes of H9N2avian influenza viruses isolated from commercial chickens in Iran[J].Trop anim health produc,2010,42(6):1291-1297.
[9]Perk S,Golender N,Banet-Noach C,et al.Phylogenetic analysis of hemagglutinin,neuraminidase,and nucleoprotein genes of H9N2avian influenza viruses isolated in Israel during the 2000-2005epizootic[J].Comp immunol,m icrobiol infect dis,2009,32(3):221-238.
[10]Tosh C,Nagarajan S,Behera P,et al.Genetic analysis of H9N2avian influenza viruses isolated from India[J].Arch Virol,2008,153(8):1433-1439.
[11]Perk S,Panshin A,Shihmanter E,et al.Ecology and molecular epidem iology of H9N2avian influenza viruses isolated in Israel during 2000-2004epizootic[J].Devel Biologicals,2006,124:201-209.
[12]Lu Jianhong,Liu Xiufan,Shao Weixing,et al.Phylogenetic analysis of eight genes of H9N2subtype influenza virus:amainland China strain possessing early isolates'genes that have been circulating[J].Virus Genes,2005,31(2):163-69.
[13]Liu Jinhua,Okazaki K,ShiWeiming,et al.Phylogenetic analysis of neuram inidase gene of H9N2influenza viruses prevalent in chickens in China during 1995-2002[J].Virus Genes,2003,27(2):197-202.
[14]Lee C W,Song C S,Lee Y J,et al.Sequence analysis of the hemagglutinin gene of H9N2Korean avian influenza viruses and assessment of the pathogenic potential of isolate MS96[J].Avian Dis,2000,44(3):527-535.
[15]王泽霖,刘岩,李建丽,等.H9N2亚型禽流感抗原性变异的研究[C].中国畜牧兽医学会禽病学分会第四十二次学术研讨会论文集.贵州贵阳,2004:395-399.
[16]荣骏弓,孙进忠,刘守川,等.鸡新城疫病毒(La Sota株)、禽流感病毒(H9亚型,HL)二联灭活疫苗免疫试验及与禽流感流行株交互免疫研究[C].中国畜牧兽医学会生物制品学分会中国微生物学会兽医微生物学专业委员会学术研讨会论文集.云南丽江,2007:531-534.
[17]姜北宇,章振华,李林,等.禽流感H9亚型流行毒株交叉免疫保护试验[J].动物医学进展,2009,30(8):29-33.
猜你喜欢
小天使·一年级语数英综合(2022年2期)2022-03-30
科学大观园(2022年2期)2022-01-23
环球时报(2018-03-19)2018-03-19
猪业科学(2017年12期)2018-01-15
中国猪业(2017年11期)2017-12-11
北方牧业(2016年15期)2016-12-17
浙江农业科学(2016年11期)2016-05-04
中国医院用药评价与分析(2015年1期)2015-08-15
医学研究杂志(2015年3期)2015-06-10
小星星·阅读100分(高年级)(2014年6期)2014-06-27
