
人Bub1基因shRNA真核表达质粒的构建及其对人卵巢癌细胞SKOV3紫杉醇敏感性的影响*
2010-05-16 02:04:34王世宣翁丹卉卢运萍
华中科技大学学报(医学版) 2010年2期
周 婷 王世宣 翁丹卉 卢运萍 马 丁
华中科技大学同济医学院附属同济医院妇产科,武汉 430030
RNA干扰(RNAi)是运用双链RNA(dsRNA)引发的转录后水平基因沉默机制[1],能特异性抑制靶基因转录,进而下调相应蛋白表达水平及功能。新近的RNAi技术是利用DNA载体直接在体内表达发卡状的siRNA(shRNA),特异性封闭相应的靶mRNA,抑制mRNA的翻译。与早期的RNA干扰技术相比,该方法的特异性和干扰效率均有较大提高,已广泛应用于各种研究领域。紫杉醇类化疗药物泰素作为全球第一种被确证的微管稳定药物,在实体瘤的治疗方面取得了巨大成功,但临床上耐药病例仍频繁发生[2]。紫杉醇通过抑制微管解聚进而阻滞细胞周期杀伤肿瘤,有研究提示这种杀伤作用有赖于具有完整功能的纺锤体检查点。纺锤体检查点是细胞周期的最后关卡[3]。纺锤体检查点功能低下时有发生,而这种功能低下与某些纺锤体蛋白的低表达有关。Bub1作为纺锤体检查点的平台蛋白,在维持纺锤体检查点的稳态等方面发挥了重要作用。目前纺锤体检查点功能是国际分子生物学研究的一个热点,但国内在该领域的研究甚少,本研究旨在运用RNAi技术,特异、稳定地抑制卵巢癌细胞Bub1 mRNA及蛋白水平表达,以对其在卵巢癌细胞紫杉醇化疗过程中的功能进行初步研究,并为进一步的机制探讨打下良好基础。
1 材料和方法
1.1 材料
pEGFP-C1空质粒为本室保存;转染试剂Lipo 2000TM购自Invitrogen公司;质粒提取试剂盒购自博大泰克,凝胶回收试剂盒购自Gibco公司;限制性内切酶购自TaKaRa公司;T4DNA连接酶购自NEB公司;四甲基偶氮唑盐(MT T)、二甲亚砜(DMSO)、碘化丙啶(PI)及紫杉醇购自德国Sigma公司,PCR引物由上海生工公司合成。抗人Bub1单克隆鼠抗购自Chemicon公司,辣根过氧化物酶标记的二抗购自美国Santa Cruz公司。
1.2 人pEGFP-Bub1-shRNA真核表达载体的构建及酶切鉴定
根据GenBank提供的Bub1基因序列,使用美国Ambio公司在线设计软件,通过基因序列比对,挑选出3条特异性寡核苷酸序列:序列1(GAGTGATCACGATT TCTAA)、序 列 2(CCAUGGGAUUGGAACCCUGTT)及序列 3(CCAGGCUGAACCCAGAGAGT T),合成相应的shRNA正义链及其互补链。以 RNAi-Ready pSI-RENDNR-DsRed-Express Vector质粒为模板,采用PCR方法扩增出带有U6启动子和shRNA序列的表达框,测序后分别用EcoRⅠ和MluⅠ双酶切,插入同样以EcoRⅠ和MluⅠ双酶切的pEGFP-C1质粒,琼脂糖凝胶电泳后回收、纯化酶切产物,后者与合成序列以T4连接酶连接3 h,转化入大肠埃希菌DH5α。筛选可疑重组阳性菌株,250 r/min摇菌过夜,小提质粒后用BamHⅠ和HindⅢ双酶切鉴定,鉴定正确者送Invitrogen公司测序,测序正确者保种,将获得的人 Bub1重组质粒命名为 pEGFPBub1-shRNA,大量提取质粒进行下一步的细胞转染。
1.3 细胞培养、转染与稳定筛选
人卵巢癌SKOV3细胞系购自ATCC,细胞以体积分数为10%FBS的McCoy's 5a培养,37℃,5%CO2,常规消化传代。3×105个/孔接种 6孔板,待细胞密度达到50%~60%时转染细胞,每孔加入8 μ L脂质体和 4.0 μ g质粒,同时设立空质粒对照组及空白对照组。48 h后加入浓度为500 mg/L的G418进行筛选,待对照细胞死亡殆尽时改用维持浓度200 mg/L,2周后挑取稳定的克隆进行鉴定,具有明显沉默效应者转入培养瓶中扩大培养。
1.4 RT-PCR检测各组细胞中Bub1基因mRNA水平的表达
收集转染48 h的各组细胞,Trizol试剂一步法提取细胞总RNA,以M-MLA逆转录酶进行逆转录,PCR扩增,同时扩增GAPDH作为内参照,以检查Bub1在转录水平的表达情况。Bub1引物序列:上 游 引物 5′-TAAACCCACAGGAGCCAGGAC-3′;下游引物 5′-CAAGCCTCAACGCCCAACT-3′;扩增条件为:95℃预变性5 min;95℃30 s,56.5℃45 s,72℃30 s,29个循环;72℃延伸10 min。GAPDH 引物 :上游引物:5′-ACGGATT TGGTCGTATTGGG-3′;下 游 引物 :5′-TGAT TT TGGAGGGATCTCGC-3′;扩增条件为:95℃预变性 5 min;95℃30 s,54℃45 s,72℃30 s,30个循环;72℃延伸10 min。PCR产物经1.5%琼脂糖凝胶电泳分离,紫外灯下观察并拍照,每个样本至少重复3次。
1.5 Western blot检测各组细胞中Bub1蛋白水平的表达
转染48 h后收集细胞,提取总蛋白,40 μ g蛋白行SDS-PAGE电泳,将蛋白质转移至硝酸纤维素膜,5%脱脂奶室温封闭2 h,一抗4℃孵育过夜,用含0.05%Tween 20的TBS缓冲液(TBST)漂洗3次,每次10 min;加入相应的碱性磷酸酶标记的二抗(1∶1 000),37℃孵育1 h,用显色底物NBT/BCIP/buffer(1∶1∶250)显色,以等量β-actin蛋白质作为对照,每个样本至少重复3次。
1.6 MTT法测定各组细胞对紫杉醇的敏感性
将经过稳定筛选、扩大培养后的各组细胞,胰酶消化后制成单细胞悬液,调整细胞密度为8×103/孔接种于96孔板,待细胞贴壁后以含不同浓度(30、90、300、1 000 nmol/L)紫杉醇的MaCoy'5a培养液200 μ L进行处理,每一浓度设3个复孔,并设不加药物和不加细胞的空白以及阴性对照组,继续培养48 h,每孔加1 mg/mL MT T 溶液100 μ L 作用4 h,弃上清液,每孔加入 DMSO 150 μ L,振荡使其充分溶解,在96孔板酶标仪上测定波长为570 nm的吸光度值(A),按如下公式计算细胞生长抑制率:细胞生长抑制率(%)=1-(实验孔A/不加药物孔A)×
100%。
1.7 流式细胞仪检测各组细胞的凋亡率
收集经紫杉醇(1 μ mol/L)作用24及 48 h的转染前后各组细胞,以冷PBS洗涤之,70%乙醇于-20℃固定过夜,洗涤,离心1 500 r/min,5 min,后加入含有 10 μ g/mL PI和0.1%RNase A 的 PBS液500 μ L,室温避光染色30 min后上流式细胞仪测定细胞凋亡率。
1.8 图像分析
RT-PCR和Western blot结果经Uvp grabit Image1软件采集,Gelworks 1D Advanced V4.01软件处理分析,计算条带的积分吸光度(IA),采用目的条带与GAPDH的IA比值表示待测样本的含量。
1.9 统计学处理
运用SPSS 12.0软件对实验数据进行统计学分析,各实验结果以±s表示,差异显著性检验采用非参数检验和方差分析,以P<0.05为差异有统计学意义。
2 结果
2.1 质粒酶切、鉴定及测序
将合成的3段退火片段分别与线性载体连接、转化后,分别挑取3~4个克隆,经扩增,提取质粒DNA,双酶切鉴定及测序,确定其中的6个克隆(2、4、5、7、11及12号克隆)插入一段大约360 bp的片段(含1个U6启动子),测序证实插入片段完全正确(图1)。

图1 人真核表达质粒pEGFP-Bub1-shRNA的酶切鉴定Fig.1 Identification of the human recombinant plasmid pEGFP-Bub1-shRNA
2.2 稳定筛选
将得到2号(序列3)、7号(序列 2)及12号(序列1)克隆进行质粒的大量提取,无菌化处理质粒后转染SKOV3细胞,G418加压筛选2周后可见有细胞克隆形成,倒置荧光显微镜下观察克隆中95%以上的细胞可见较强的绿色荧光(图2),挑取克隆进行扩大培养。

图2 pEGFP-C1和pEGFP-Bub1-shRNA转染SKOV3细胞G418筛选后稳定表达(×40)Fig.2 The green fluorescence in SKOV3 cells observed under the fluorescent microscope after transfected with plasmids(×40)
2.3 各组细胞中Bub1 mRNA的表达
凝胶电泳分析各组细胞Bub1 mRNA表达的差异,由图3可见,pEGFP-C1/SKOV3组和SKOV3组细胞约450 bp处条带清晰明亮,7号及12号克隆所对应的pEGFP-Bub1-shRNA/SKOV3在同样位置出现模糊条带,而2号克隆与对照组相比无明显差异。经相对强度对比分析,7号及12号克隆相对表达强度分别为(0.25±0.05)和(0.18±0.03),pEGFP-C1/SKOV3组为(0.70±0.08),7号及12号克隆与pEGFP-C1/SKOV3组相比差异均有显著性意义(P<0.05),12号克隆的干扰效应强于7号克隆。以上结果说明,序列1对应的pEGFP-Bub1-shRNA质粒的干扰效应在三者中最强,后续实验均以该质粒为基础进行。

图3 各组细胞中Bub1 mRNA的表达Fig.3 Bub1 mRNA expression in cells of each group
2.4 各组细胞蛋白质水平的表达
由图4可见,pEGFP-Bub1-shRNA/SKOV3细胞Bub1蛋白相对表达强度(Bub1/β-actin)为(0.10±0.04),而 pEGFP-C1/SKOV3和 SKOV3细胞Bub1蛋白相对表达强度分别为(0.80±0.03)和(0.78±0.04),以上结果说明,pEGFP-Bub1-shRNA/SKOV3细胞Bub1蛋白表达水平较 pEGFPC1/SKOV3和SKOV3两组细胞明显降低,差异具有统计学意义(P<0.05)。

图4 各组中Bub1蛋白的表达Fig.4 Bub1 protein expression in cells of each group
2.5 各组SK OV3细胞紫杉醇敏感性
如图5所示,紫杉醇浓度为 90、300及1 000 nmol/L时,pEGFP-Bub1-shRNA/SKOV3组细胞的生长抑制率显著低于pEGFP-C1/SKOV3和SKOV3细胞,差异均具有统计学意义(P<0.05);由表1可以看到,以1 μ mol/L紫杉醇作用 24及 48 h,pEGFP-Bub1-shRNA/SKOV3组细胞的凋亡率显著低于pEGFP-C1/SKOV3组,差异具有统计学意义(P<0.01);pEGFP-C1/SKOV3和 SKOV3细胞的生长抑制率和凋亡率相比差异均无统计学意义。
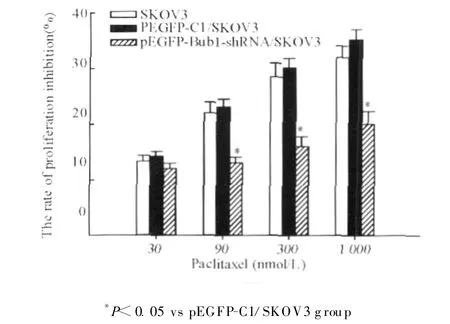
图5 各组细胞的生长抑制率Fig.5 The inhibition rate of proliferation in each group
表1 1μ mol/L紫杉醇处理后不同时间点各组细胞凋亡率(±s,%)Table 1 The apoptosis rate of cells treated with paclitaxel at different time points in each g roup(±s,%)

表1 1μ mol/L紫杉醇处理后不同时间点各组细胞凋亡率(±s,%)Table 1 The apoptosis rate of cells treated with paclitaxel at different time points in each g roup(±s,%)
*P<0.01 vs pEGFP-C1/SKOV3 group
Groups 0 h 24 h 48 h SKOV3 1.63±0.35 10.14±1.73 34.17±3.22 pEGFP-C1/SKOV3 2.43±0.42 12.15±1.97 37.22±3.88 pEGFP-Bub1-shRNA/SKOV3 2.20±0.27 4.42±0.62*12.98±1.94*
3 讨论
RNAi现象自1998年被发现以来,已逐步成为肿瘤相关基因领域的研究热点。从小分子干扰RNA技术到shRNA技术,RNAi技术在稳定性和临床应用潜能上得到了深入发展[4]。siRNA干扰效果不稳定,在哺乳动物中时效性差,且siRNA在细胞内容易被降解,对 RNase-free环境要求高,与siRNA技术相比,shRNA技术既保持了siRNA特异性高的优势,且其具有更强的稳定性,能够持续发挥抑制作用以便研究者在更长的时间跨度中对基因功能进行深入探索。本研究构建的shRNA真核表达质粒pEGFP-Bub1-shRNA转染人卵巢癌细胞SKOV3后,该细胞内Bub1 mRNA及蛋白表达水平明显下调,证实构建的pEGFP-Bub1-shRNA真核表达质粒能高效阻断Bub1基因表达,为进一步研究Bub1基因的生物学功能提供了有力工具。
细胞在长期的进化过程中产生的监控纺锤体形态、染色体着丝点与微管联接及其产生的张力、染色体排列、确保姐妹染色单体精确分离的质控机制,即纺锤体检查点,又称有丝分裂检查点[5]。蛋白激酶Bub1作为纺锤体检查点的平台蛋白,可能是纺锤体检查点其他组分定位于纺锤体检查点的基础。Bub1可通过直接作用及形成复合物的形式来调控其他重要的纺锤体检查点蛋白,如Bub3、Mad2及CDC20等,进而影响细胞周期的进程[6]。虽然临床大样本的研究提示纺锤体检查点基因突变频率极低,但多种肿瘤细胞确实存在纺锤体检查点功能缺陷,而有研究表明这种缺陷可能与纺锤体检查点蛋白表达异常有关。有文献报道急性髓细胞白血病细胞中存在Bub1 mRNA低表达[7];Shichiri及其研究小组[8]发现大肠癌组织Bub1整体表达水平明显高于邻近正常组织,与此同时,少数Bub1 mRNA表达水平明显低于正常组织的大肠癌似乎更易于发生淋巴结转移和复发;此外,有研究发现,胃癌组织Bub1 mRNA显著高表达,且与癌细胞增殖相关[9]。早在提出“纺锤体检查点”概念前,微管稳定剂之一的紫杉醇类药物就已广泛应用于临床,它们的主要作用靶点就是纺锤体检查点。虽然这类药物在临床应用上取得了巨大成功,但耐药现象仍时有发生。相关研究指出紫杉醇类药物的敏感性必须依赖于功能完整的纺锤体检查点[10],因此,若想提高这些抗癌药物的效能,就必须对纺锤体检查点蛋白的功能进行更为深入的研究。目前,该领域的研究是国际癌症研究的热点之一,国内研究尚处于起步阶段,文献报道极少。
我们运用RNAi技术在转录后水平特异性干扰SKOV3细胞中Bub1 mRNA的表达,进而降低了Bub1蛋白表达水平,随后经紫杉醇处理,发现肿瘤细胞生长抑制率显著下降,凋亡细胞明显减少,由此初步证明Bub1在卵巢癌细胞化疗耐药中发挥了重要作用。有研究者推测纺锤体检查点功能缺陷的细胞在纺锤体抑制因子持续或高剂量作用后,可不停滞于过渡期而变为多倍体细胞,继续存活;正常细胞则长期停滞于该期,因纺锤体未及时修复而发生凋亡。我们在SKOV3细胞模型中的实验结果为上述研究提供了佐证。尽管如此,Bub1表达低下时哪些关键下游基因随之变化,Bub1基因的下调是否直接影响纺锤体检查点的功能,纺锤体检查点的失活是否导致肿瘤细胞化疗耐药,这些问题还有待于深入探讨。值得注意的是,在不同的细胞系,纺锤体检查点的功能呈现多元化的趋势,例如,我们的前期研究提示,在某些宫颈癌细胞株中,通过蛋白抑制剂破坏其纺锤体检查点蛋白Bub1可增强紫杉醇的杀灭效能,故在这类细胞中运用pEGFP-Bub1-shRNA载体抑制Bub1的表达亦可能增强宫颈癌辅助化疗的敏感性。综上所述,深入了解纺锤体检查点蛋白功能对于探讨纺锤体检查点的作用机制和提高肿瘤细胞化疗疗效均具有十分重要的意义,而我们前期的相关工作为后续研究奠定了良好的基础。
[1] Hammond S M,Emily B,David B,et al.An RNA-directed nuclease mediates post-transcriptional gene silencing in Drosophila cells[J].Nature,2000,404(6775):293-295.
[2] Berg stralh D T,Ting J P.Microtubule stabilizing agents:their molecular signaling consequences and the potential for enhancement by drug combination[J].Cancer T reat Rev,2006,32(3):166-179.
[3] 傅云峰,谢幸,叶大风.肿瘤与纺锤体检查点的研究现状[J].国外医学·肿瘤学分册,2005,32(1):3-6.
[4] 李力,张会强,贺福初.RNA干拢应用新展[J].医学分子生物学杂志,2004,1(4):224-226.
[5] Wassmann K,Benezra R.Mitotic checkpoints:from yeast to cancer[J].Curr Opin Genet Dev,2001,11(1):83-90.
[6] Bharadwaj R,Yu H.T he spindle checkpoint,aneuploidy,and cancer[J].Oncogene,2004,23(11):2016-2027.
[7] Lin S F,Lin P M,Yang M C,et al.Expression of hBUB1 in acute my eloid leukemia[J].Leuk Lymphoma,2002,43(2):385-391.
[8] Shichiri M,Yoshinaga K,Hisatomi H,et al.Genetic and epigenetic inactivation of mitotic checkpoint genes hBUB1 and hBUBR1 and their relationship to survival[J].Cancer Res,2002,62(1):13-17.
[9] Grabsch H,Takeno S,Parsons W J,et al.Overex pression of the mitotic checkpoint genes BUB1,BUBR1,and BUB3 in gastric cancer-association with tumour cell proliferation[J].J Pathol,2003,200(1):16-22.
[10] Sudo T,Nitta M,Saya H,et al.Dependence of paclitaxel sensitivity on a functional spindle assembly checkpoint[J].Cancer Res,2004,64(7):2502-2508.
猜你喜欢
中国医学科学院学报(2022年1期)2022-12-30 13:30:21
计算机系统应用(2022年4期)2022-05-10 08:41:10
天津医科大学学报(2021年4期)2021-08-21 02:14:52
诊断学(理论与实践)(2020年2期)2020-06-28 12:41:12
国际呼吸杂志(2019年4期)2019-03-12 01:08:18
实用口腔医学杂志(2017年6期)2017-09-19 02:51:06
生物学教学(2017年9期)2017-08-20 13:22:32
中外医疗(2016年15期)2016-12-01 04:25:50
哈尔滨医药(2015年2期)2015-12-01 03:57:41
现代计算机(2015年31期)2015-09-28 05:31:51
