
基于YTHDF2 介导凋亡相关因子降解途径研究牙龈卟啉单胞菌协助食管癌免疫逃逸
2024-11-03 00:00:00杨泽张秀森张旭东柳颖张嘉诚原翔
南方医科大学学报 2024年6期


摘要:目的 探讨牙龈卟啉单胞菌(Pg)感染食管癌细胞后对YTHDF2 及凋亡相关因子(Fas)的影响,阐明其促进免疫逃逸可能的调控机制。方法 运用免疫组织化学法及Western blot法检测Pg感染食管癌与YTHDF2及Fas表达变化;运用免疫共沉淀验证YTHDF2和Fas蛋白间的相互作用;将体外培养的Pg感染后的KYSE150细胞通过慢病毒转染分为对照组(转染si-NC)和实验组(转染si-YTHDF2),Western blotting 检测两组细胞中YTHDF2、组织蛋白酶B(CTSB)及Fas、FasL蛋白的表达水平;对Pg感染的KYSE150细胞用组织蛋白酶抑制剂(E64)进行处理,Western blotting分别检测抑制剂处理前后YTHDF2、CTSB、Fas及FasL蛋白的表达变化情况;Pg感染前后及E64处理的KYSE150细胞与人外周血单个核细胞(PBMC)共培养,运用流式细胞术检测T细胞相关效应分子的表达情况。结果 免疫组化法及Western blotting 结果显示,Pg 感染后的组织或细胞中YTHDF2 的表达增强,而Fas 表达减弱(Plt;0.001);免疫共沉淀结果显示,YTHDF2 和Fas 之间可直接相互作用;Western blotting 结果显示,si-YTHDF2组细胞中CTSB表达量减低,而Fas及FasL的表达量高于si-NC组(Plt;0.001);用E64处理细胞后,CTSB表达减弱,YTHDF2表达无影响,而Fas及FasL的表达增强;流式细胞术结果显示,与PBMC共培养的细胞中,GranzymeB及Ki67表达由强到弱分别为Pg 阴性、Pg 阳性+E64、Pg 阳性,而PD-1 表达情况相反(Plt;0.001)。结论 Pg 感染依赖YTHDF2 调节Fas的表达,协助食管癌免疫逃逸,从而促进食管癌发生发展,表明YTHDF2可能是一个新的调控食管癌免疫逃逸的关键分子。
关键词:食管鳞状细胞癌;牙龈卟啉单胞菌;m6A甲基化阅读蛋白YTHDF2;凋亡相关因子;肿瘤免疫
食管癌是最常见且最致命的恶性肿瘤之一,是全球癌症相关死亡的第6大原因[1]。食管癌早期缺乏特异性症状或体征,因此总体预后极差,5 年总生存率仅15%左右。食管癌按照组织学类型主要分为鳞状细胞癌(ESCC)和腺癌,其中ESCC约占80%以上[2]。食管癌的病因和发病机制可能与饮食、地域、遗传、感染等多种因素相关[3, 4],我国ESCC的发生发展可能具有其独特的高危因素及分子机制[5, 6],但目前尚未被完全阐明。因此,需要进一步对食管癌相关的分子机制进行深入研究,探索可能存在的新型特异性生物标志物,为食管癌的诊治提供新策略。
人体消化道存在多种微生物定植,以细菌为主,维持消化道微生物菌群平衡对消化道健康有重要作用。牙龈卟啉单胞菌(Pg)作为牙周炎的标志性病原微生物,其特性包括:破坏口腔微环境稳态、调控细胞增殖或凋亡、激活上皮细胞的转化、促进血管生成、抑制宿主免疫系统以及产生致癌代谢物等[7]。有研究表明,Pg在食管中定植并且Pg感染与食管癌的发生发展密切相关;Pg感染能够通过FimA等毒力因子通过多种途径协助肿瘤细胞免疫逃逸,进一步促进食管癌细胞增殖、迁移、侵袭和转移能力[8, 9]。
N6-甲基腺苷(m6A)为真核生物mRNA最常见、最丰富的转录后修饰,参与RNA的剪接、转录、翻译和降解等各种代谢过程[10]。研究表明,异常的m6A甲基化及其关键蛋白的表达与多种恶性肿瘤发生和进展过程密切相关[11]。YTHDF为甲基阅读器蛋白,可与m6A特异性结合选择性调节RNA的代谢[12]。YTH N6-甲基腺苷RNA结合蛋白2(YTHDF2)是m6A修饰的关键性蛋白,已在多种恶性肿瘤的文献中报道[13]。研究发现,m6A甲基化阅读蛋白YTHDF2能够调控肿瘤相关巨噬细胞的抗肿瘤免疫,且在CD8+T细胞的参与下,YTHDF2显著降低了肿瘤细胞的先天性和适应性免疫[14]。有研究表明,Pg能够通过RNA甲基化等表观遗传修饰途径促进多种口腔炎症性疾病的进展[15]。本团队前期研究表明,Pg感染可促进食管癌细胞逃避宿主的免疫监视[9],然而Pg感染后肿瘤细胞逃避免疫的能力是否与m6A甲基化阅读蛋白相关目前相关尚未有报道。因此Pg感染通过m6A甲基化阅读蛋白YTHDF2途径影响肿瘤免疫的相关机制有待进一步阐明。
凋亡相关因子(Fas)属于肿瘤坏死因子受体超家族,是一种死亡受体,与配体FasL结合后能够介导细胞的凋亡,在维持免疫微环境稳态等过程中发挥重要作用[16]。Fas的异常表达与多种免疫相关性肿瘤和疾病的发病机制存在密切联系,有研究发现抑制癌症细胞中Fas的表达能够诱导CD8+T细胞凋亡,降低肿瘤细胞对获得性免疫反应的敏感性[17]。FasL高表达的肿瘤细胞可以通过Fas/FasL相互作用来传递凋亡信号,促进肿瘤浸润淋巴细胞的凋亡,最终导致抗肿瘤免疫逃逸[18]。推测在Pg感染的食管癌中可能也存在这种情况。
本研究通过检测Pg 感染食管癌后对YTHDF2 的影响,以及YTHDF2与Fas之间的相互作用,阐明Pg感染对CD8+T细胞免疫功能的抑制作用,初步探索Pg感染与甲基化阅读蛋白YTHDF2的相关机制。
1 材料和方法
1.1 实验材料及主要试剂
1.1.1 组织样本
纳入2022 年9 月~2023 年5 月在我院被诊断为食管鳞癌并进行手术的患者病理组织。本研究经医院伦理委员会的批准(伦理审批编号:2021-0165),患者均签署知情同意书。术中切除的肿瘤组织用多聚甲醛固定后蜡块包埋以备用。
1.1.2 实验主要试剂
ATCC 33277 来源于本实验室;KYSE150细胞系(上海博古生物细胞所);细胞培养基、胎牛血清(FBS)、胰蛋白酶、青-链霉素、免疫组织化学染色试剂盒、PBS磷酸盐缓冲液(北京索莱宝科技有限公司);PAGE凝胶制备试剂盒(10%)、e-ECL显影液(雅酶公司);Anti-YTHDF2 抗体、Anti-CTSB 抗体(ThermoFisher Scientific);Anti-Fas 抗体、Anti-FasL 抗体(Abcam);Pg 抗体diatheva(上海玉博生物科技有限公司);PVDF膜(milipore Milipore);倒置显微镜(Nikon);全自动酶标仪、细胞培养箱(Thermo);凝胶成像分析仪(Bio-Rad);荧光共聚焦显微镜(ZEISS);厌氧培养箱(Shellab);流式细胞仪(CyTek)。
1.2 实验方法
1.2.1 细胞培养及Pg培养
KYSE150细胞培养:含10%胎牛血清和1%双抗的RPMI 1640完全培养基,放置于含有5% CO2的37 ℃恒温培养箱中培养,必要时进行传代或冻存处理。Pg标准菌株ATCC 33277于37 ℃厌氧条件下培养,每次感染细胞前,采用革兰染色后进行形态学检测及16S rDNA特异扩增检查确定菌株未发生变异及其它细菌污染。
1.2.2 Pg感染细胞
待被感染细胞生长至80%以上时,PBS清洗细胞2次,加入1 mL胰蛋白酶置于恒温培养箱中消化5 min,加入2 mL RPMI 1640 培养基终止消化。消化后的细胞悬液离心、弃上清、重悬细胞。在细胞计数后向新的培养皿内加入(1~10)×106 KYSE150细胞,于恒温培养箱中培养细胞至完全贴壁,弃去原细胞培养液,用PBS清洗细胞,加入RPMI 1640培养基。取生长期细菌感染细胞,培养Pg使其OD值达1.0~1.5,将Pg悬液以12 000 r/min 转速离心10 min 后用RPMI 1640 重悬。按照预定的感染复数值在细胞培养基中加入相应数量的Pg,随后于恒温培养箱中培养3 d。3 d后收集细胞培养液并于4000 r /min条件下离心20 min,留取上清进一步使用0.22 μm孔径的滤器过滤,滤液置于-20 ℃冰箱保存。将体外培养的Pg 感染后的KYSE150 细胞通过慢病毒转染分为对照组(转染si-NC)和实验组(转染si-YTHDF2)。
1.2.3 免疫组织化学 (IHC)
将包埋好的蜡块切片、烤片。依次将载玻片放入二甲苯、梯度酒精中各3 min脱蜡,3% H2O2中浸泡10 min,在清水中洗2 次,再置入柠檬酸缓冲液中,放入微波炉煮至沸腾,取出载玻片冷却至室温。滴加血清封闭液孵育30 min,加入适量一抗4 ℃孵育过夜,室温孵育二抗。加显色剂,用清水冲洗一段时间后,浸泡于苏木精中染色。载玻片依次放入梯度酒精、二甲苯中脱水,晾干后中性树胶封片。
1.2.4 Western blotting
配制细胞裂解液裂解后提取蛋白,取少量样品用BCA法测蛋白浓度,其余蛋白按照测定浓度加入Loading Buffer,95 ℃煮10 min。制备浓缩胶,等质量蛋白上样后进行电泳,转移到PVDF 膜上,5%脱脂奶粉封闭1 h,TBST洗膜。根据实验需要一抗4 ℃孵育过夜,TBST洗膜,室温孵育二抗1 h,TBST洗膜。ECL方法进行曝光显影。
1.2.5 免疫共沉淀 (Co-IP)
在293T 细胞中,用Myc-YTHDF2 和编码HA-Fas 的表达质粒进行转染。细胞加入适量RIPA细胞裂解缓冲液,置于冰上30 min,4 ℃最大转速离心30 min后取上清。取少量裂解液以备分析(Input),剩余裂解液分别使用Myc和HA抗体与磁珠结合后共同孵育得到免疫沉淀复合物。剩余裂解液中加入1 μg 相应抗体制成混合液,4 ℃缓慢摇晃孵育过夜,混合液中再加入10 μL 预处理(裂解缓冲液洗3次,每次3000 r/min,离心3 min)过的protein A 琼脂糖珠,摇晃孵育2 h。4 ℃ 3000 r/min离心3 min,吸去上清,裂解缓冲液洗3次,加入SDS加样缓冲液,煮沸5 min。最终所得样品和Input均用Western blotting检测。
1.2.6 流式细胞术
人外周血和1640培养基按照1∶1的比例加入离心管内,分层加入适量淋巴细胞分离培养基(FicollPaque),500 g 离心25 min,吸取中间的白色薄膜层,即为PBMC。PBS洗涤2次,调整细胞密度至1×106/mL。与KYSE150细胞按2∶1的比例共培养48 h后再次提取PBMC。加入适量细胞染色缓冲液重悬细胞并转移至流式管,加入流式抗体,4 ℃避光孵育30 min,细胞染色缓冲液洗涤,离心弃上清,再加入适量细胞染色缓冲液重悬细胞,上机进行检测。
1.3 统计学方法
使用SPSS 26.0 软件进行统计分析,使用GraphPad Prism 7软件绘图。实验数据以均数±标准差表示,两组间比较采用t检验,多组间比较采用单因素方差分析。Plt;0.05为差异具有统计学意义。
2 结果
2.1 食管鳞癌中Pg感染与YTHDF2和Fas蛋白表达变化的相关性
牙龈卟啉单胞菌感染ESCC组织的免疫组化结果显示Pg 在肿瘤细胞的胞浆染色(图1A、B)。免疫组化结果显示,与Pg 阴性的ESCC的组织相比,Pg 阳性的组织中YTHDF2 的表达量增强,而Fas 的表达量减弱(图1C~F,Plt;0.001)。Western blot结果显示,Pg阳性的KYSE150细胞内YTHDF2的表达量增加,相反的,Fas的表达量减弱(图1G~I,Plt;0.001)。
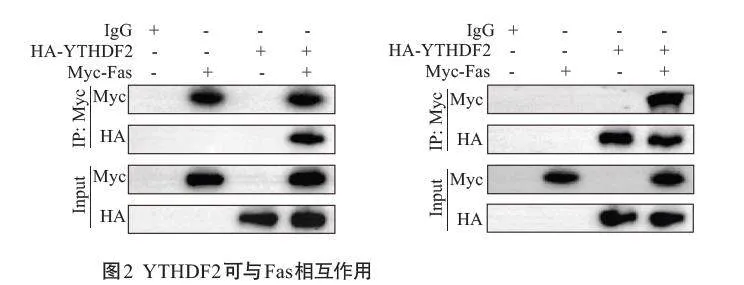
2.2 YTHDF2可与Fas相互作用
Co-IP实验结果显示,外源性YTHDF2能够与外源性Fas相互结合形成复合物(图2)。
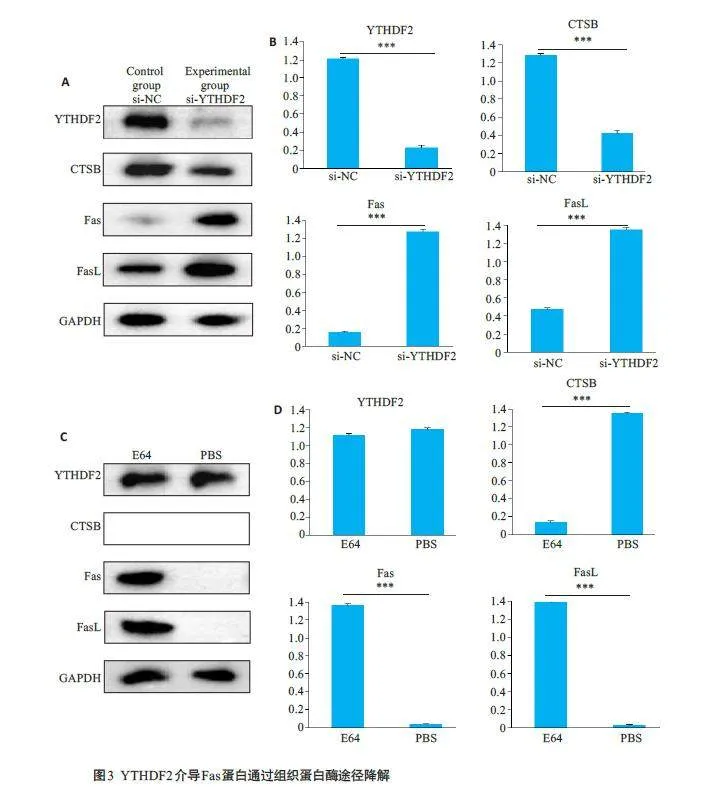
2.3 YTHDF2介导Fas蛋白通过组织蛋白酶途径降解
Western blotting结果显示,相比对照组,实验组中YTHDF2 蛋白的表达量减少,CTSB的表达量也减少,而Fas、FasL 的表达量增加(图3A、B,Plt;0.001)。Western blot结果显示,在E64处理后,YTHDF2表达量不变,CTSB 表达量减弱,而Fas 和FasL 表达量增强(图3C、D,Plt;0.001)。
2.4 Pg 依赖YTHDF2-Fas 途径抑制食管癌中T细胞的免疫效应
流式细胞术结果显示,Pg 感染组的Granzyme B,Ki-67 的表达减弱,PD-1 表达增强(Plt;0.001),但是经E64 处理可部分恢复Granzyme B和Ki-67 的表达水平(图4)。
3 讨论
病原微生物具有复杂多样的生物功能,是肿瘤内在微环境的重要组成部分[19]。大多数病原微生物并不能直接导致癌症,但它们通常会影响宿主正常的免疫功能,协助参与癌症的发生发展过程[20]。牙龈卟啉单胞菌被认为是慢性牙周炎的关键病原体,可导致微环境失调和异常的免疫反应[21]。有研究表明,牙龈卟啉单胞菌能够通过PGN诱导PD-L1 上调,改变肿瘤免疫微环境来促进前列腺癌的发展和进展[22];而在结直肠癌中,牙龈卟啉单胞菌能够侵入肿瘤细胞并激活MAPK/ERK信号通路,从而促进结直肠癌细胞的增殖[23]。Pg感染可导致多种恶性肿瘤肿瘤进展,但目前没有研究阐述牙龈卟啉单胞菌与m6A甲基化的相关机制。本研究主要从Pg感染通过m6A 甲基化阅读蛋白YTHDF2 途径协助ESCC细胞免疫逃避,进而促进食管癌进展这一角度阐明相关机制。
虽然已有研究表明,牙龈卟啉单胞菌可能通过miR-194/GRHL3/PTEN/Akt 信号轴促进ESCC的增殖和迁移[24];也可能通过促进IFNGR1在122位点的棕榈酰化下调IFNGR1的蛋白表达,从而增强ESCC细胞增殖、迁移和侵袭[25] ;或者通过GSK3β 介导的mtOXPHOS促进ESCC的进展[26]。研究表明YTHDF2作为m6A甲基化的关键阅读蛋白,通过调控mRNA的翻译及降解等导致m6A甲基化水平发生改变,从而影响下游分子的表达水平[27]。本研究发现,Pg 阳性的ESCC肿瘤组织或细胞中,YTHDF2表达增高。这表明Pg可能通过上调YTHDF2介导其依赖m6A甲基化修饰途径来促进ESCC的进展,需要进一步研究Pg 感染能否通过甲基化阅读蛋白影响m6A甲基化水平的变化。
多种肿瘤细胞表面的Fas 表达水平减低,通过与配体间的相互作用,诱导细胞凋亡,进一步逃避免疫系统监视,但其潜在的机制尚不明确[28]。本研究发现YTHDF2与Fas能直接相互作用,并且YTHDF2高表达能够诱导Fas 降解。我们发现Pg 感染ESCC 后YTHDF2高表达,而Fas低表达。CTSB是一种定位于溶酶体的半胱氨酸蛋白酶,CTSB与细胞中溶酶体和自噬相关蛋白的表达密切相关[29]。因此,我们猜测这二者呈现负相关的原因可能是YTHDF2 诱导自噬途径增强。自噬途径是将各种细胞代谢产物输送到溶酶体进行降解或再循环的主要机制[30]。有研究显示,YTHDF2通过调控体细胞重编程介导mRNA降解,促进体细胞相关基因的自噬清除[31],证明了YTHDF2 能够调控mRNA代谢过程,促进细胞代谢分子通过自噬途径降解。本研究通过实验进一步证明了Pg 感染高表达YTHDF2能够介导Fas通过自噬途径降解。此外,我们还利用肿瘤细胞和PBMC共培养,通过流式细胞术检测证明Pg感染ESCC后可以显著减弱T细胞的增殖和杀伤作用,促进食管癌免疫逃避。
牙龈卟啉单胞菌可释放多种毒力因子,包括菌毛、荚膜、蛋白酶和外膜囊泡等,这些毒力因子的产生能够使细菌更好地在宿主体内存活、繁殖,并且其致病性在很大程度上与干扰宿主的免疫功能有关[32, 33]。菌毛蛋白FimA是牙龈卟啉单胞菌的重要毒力因子,在促进细菌黏附和入侵宿主细胞、参与生物膜形成以及引起宿主细胞的免疫炎症反应等多种过程中发挥重要作用[34]。有研究表明,Pg感染后其毒力因子FimA的基因表达在上消化道肿瘤中呈现上高下低的分布特征[35]。有研究显示,FimA缺失可能降低菌体间黏附,使侵入细胞内的细菌数目减少,刺激肿瘤细胞增殖、迁移和侵袭能力降低;这进一步证明可能存在FimA 介导Pg 从而促进ESCC进展的相关机制,是Pg促肿瘤作用的潜在分子靶点[36]。因此,我们推测Pg在调控YTHDF2高表达的过程中,可能通过FimA等致病毒力因子,这需要进一步研究证实。
综上所述,本研究初步提出Pg感染可能通过m6A甲基化阅读蛋白YTHDF2途径介导Fas降解,并进一步抑制T细胞杀伤能力,协助食管癌产生免疫逃逸。
参考文献:
[1] Sung H, Ferlay J, Siegel RL, et al. Global cancer statistics 2020:GLOBOCAN estimates of incidence and mortality worldwide for 36cancers in 185 countries[J]. CA Cancer J Clin, 2021, 71(3): 209-49.
[2] Pennathur A, Gibson MK, Jobe BA, et al. Oesophageal carcinoma[J]. Lancet, 2013, 381(9864): 400-12.
[3] Zhou JC, Sun KX, Wang SM, et al. Associations between cancer family history and esophageal cancer and precancerous lesions inhigh-risk areas of China[J]. Chin Med J, 2022, 135(7): 813-9.
[4] Xie SH, Lagergren J. Risk factors for oesophageal cancer[J]. BestPract Res Clin Gastroenterol, 2018, 36/37: 3-8.
[5] Cancer Genome Atlas Research Network, Analysis Working Group:Asan University, BC Cancer Agency, et al. Integrated genomiccharacterization of oesophageal carcinoma[J]. Nature, 2017, 541(7636): 169-75.
[6] Deng JY, Chen H, Zhou DZ, et al. Comparative genomic analysis ofesophageal squamous cell carcinoma between Asian and Caucasianpatient populations[J]. Nat Commun, 2017, 8(1): 1533.
[7] Lamont RJ, Fitzsimonds ZR, Wang HZ, et al. Role ofPorphyromonas gingivalis in oral and orodigestive squamous cellcarcinoma[J]. Periodontol 2000, 2022, 89(1): 154-65.
[8] Gao SG, Yang JQ, Ma ZK, et al. Preoperative serumimmunoglobulin G and A antibodies to Porphyromonas gingivalisare potential serum biomarkers for the diagnosis and prognosis ofesophageal squamous cell carcinoma[J]. BMC Cancer, 2018, 18(1): 17.
[9] Yuan X, Liu YW, Li GF, et al. Blockade of immune-checkpoint B7-H4 and lysine demethylase 5B in esophageal squamous cellcarcinoma confers protective immunity against P. gingivalisinfection[J]. Cancer Immunol Res, 2019, 7(9): 1440-56.
[10]Zhuang HZ, Yu B, Tao D, et al. The role of m6A methylation intherapy resistance in cancer[J]. Mol Cancer, 2023, 22(1): 91.
[11] Lan Q, Liu PY, Haase J, et al. The critical role of RNA m6Amethylation in cancer[J]. Cancer Res, 2019, 79(7): 1285-92.
[12]Zaccara S, Jaffrey SR. A Unified Model for the Function of YTHDFProteins in Regulating m(6)A-Modified mRNA[J]. Cell, 2020, 181(7): 1582-95.
[13]Wang X, Zhao B, Roundtree I, et al. N6-methyladenosine modulatesmessenger RNA translation efficiency[J]. Cell, 2015, 161(6):1388-99.
[14]Ma SB, Sun BF, Duan SQ, et al. YTHDF2 orchestrates tumorassociatedmacrophage reprogramming and controls antitumorimmunity through CD8+ T cells[J]. Nat Immunol, 2023, 24(2):255-66.
[15]Huang J, Zhou Y. Emerging role of epigenetic regulations inperiodontitis: a literature review[J]. Am J Transl Res, 2022, 14(4):2162-83.
[16]Lagunas-Rangel FA. Fas (CD95)/FasL (CD178) system duringageing[J]. Cell Biol Int, 2023, 47(8): 1295-313.
[17]Mahadevan KK, McAndrews KM, LeBleu VS, et al. KRAS(G12D)inhibition reprograms the microenvironment of early and advancedpancreatic cancer to promote FAS-mediated killing by CD8+ T cells[J]. Cancer Cell, 2023, 41(9): 1606-20.
[18]Chiu HY, Sun GH, Chen SY, et al. Pre-existing fas ligand (FasL) incancer cells elicits tumor-specific protective immunty, but delayedinduction of FasL expression after inoculation facilitates tumorformation[J]. Mol Carcinog, 2013, 52(9): 705-14.
[19]Xie YF, Xie F, Zhou XX, et al. Microbiota in tumors: fromunderstanding to application[J]. Adv Sci, 2022, 9(21): e2200470.
[20]Sepich-Poore GD, Zitvogel L, Straussman R, et al. The microbiomeand human cancer[J]. Science, 2021, 371(6536): eabc4552.
[21]Singhrao SK, Harding A, Simmons T, et al. Oral inflammation, toothloss, risk factors, and association with progression of Alzheimer'sdisease[J]. J Alzheimers Dis, 2014, 42(3): 723-37.
[22]Groeger S, Wu F, Wagenlehner F, et al. PD-L1 up-regulation inprostate cancer cells by Porphyromonas gingivalis[J]. Front CellInfect Microbiol, 2022, 12: 935806.
[23]Mu WX, Jia YQ, Chen XB, et al. Intracellular Porphyromonasgingivalis promotes the proliferation of colorectal cancer cells viathe MAPK/ERK signaling pathway[J]. Front Cell Infect Microbiol,2020, 10: 584798.
[24]Liang GF, Wang HJ, Shi H, et al. Porphyromonas gingivalispromotes the proliferation and migration of esophageal squamouscell carcinoma through the miR-194/GRHL3/PTEN/akt axis[J].ACS Infect Dis, 2020, 6(5): 871-81.
[25]Shen L, Zhang D, Gao S. Effect of infection on IFNGR1palmitoylation in esophageal cancer cells [J]. J South Med Uni,2023, 43: 1155-63.
[26]Liu YW, Zhou FY, Yang HJ, et al. Porphyromonas gingivalispromotes malignancy and chemo-resistance via GSK3β‑mediatedmitochondrial oxidative phosphorylation in human esophagealsquamous cell carcinoma[J]. Transl Oncol, 2023, 32: 101656.
[27]Dixit D, Prager BC, Gimple RC, et al. The RNA m6A readerYTHDF2 maintains oncogene expression and is a targetabledependency in glioblastoma stem cells[J]. Cancer Discov, 2021, 11(2): 480-99.
[28]Sharma S, Carmona A, Skowronek A, et al. Apoptotic signallingtargets the post-endocytic sorting machinery of the death receptorFas/CD95[J]. Nat Commun, 2019, 10(1): 3105.
[29]Man SM, Kanneganti TD. Regulation of lysosomal dynamics andautophagy by CTSB/cathepsin B[J]. Autophagy, 2016, 12(12):2504-5.
[30]Debnath J, Gammoh N, Ryan KM. Autophagy and autophagyrelatedpathways in cancer[J]. Nat Rev Mol Cell Biol, 2023, 24(8):560-75.
[31]Liu JD, Gao MW, Xu SY, et al. YTHDF2/3 are required for somaticreprogramming through different RNA deadenylation pathways[J].Cell Rep, 2020, 32(10): 108120.
[32]Zheng SW, Yu SW, Fan XM, et al. Porphyromonas gingivalissurvival skills: immune evasion[J]. J Periodontal Res, 2021, 56(6):1007-18.
[33]Li C, Yu R, Ding YM. Association between PorphyromonasGingivalis and systemic diseases: focus on T cells-mediatedadaptive immunity[J]. Front Cell Infect Microbiol, 2022, 12:1026457.
[34]Xu WZ, Zhou W, Wang HZ, et al. Roles of Porphyromonasgingivalis and its virulence factors in periodontitis[J]. Adv ProteinChem Struct Biol, 2020, 120: 45-84.
[35]Yuan X, Liu YW, Kong JY, et al. Different frequencies ofPorphyromonas gingivalis infection in cancers of the upperdigestive tract[J]. Cancer Lett, 2017, 404: 1-7.
[36]阮豪杰, 程维刚, 焦叶林, 等. 菌毛素介导牙龈卟啉单胞菌对食管鳞癌促进作用[J]. 中华微生物学和免疫学杂志, 2022, 42(4): 275-80.
(编辑:林 萍)
基金项目:河南省科学技术厅-优秀青年科学基金(222300420041);河南省教育厅-河南省科技攻关项目(242102310146)
猜你喜欢
中国科技纵横(2021年24期)2021-03-02 06:42:52
中国生殖健康(2019年6期)2019-01-06 09:20:46
中成药(2018年6期)2018-07-11 03:01:12
知识就是力量(2018年6期)2018-06-15 07:32:58
小学生导刊(2018年4期)2018-04-18 13:53:31
中成药(2017年9期)2017-12-19 13:34:21
中华老年口腔医学杂志(2016年2期)2017-01-15 14:24:48
现代检验医学杂志(2015年1期)2015-02-06 01:59:05
食品工业科技(2014年23期)2014-03-11 18:19:08
山西大同大学学报(自然科学版)(2013年5期)2013-09-13 10:44:14
