
锌指蛋白281抑制高糖诱导的肾小管上皮细胞上皮间质转化和细胞外基质合成
2024-07-13 09:47侯维玲乔云阳吴小芸施会敏曲高婷张爱青
天津医药 2024年7期
侯维玲 乔云阳 吴小芸 施会敏 曲高婷 张爱青

基金项目:江苏省医学会儿科医学科研专项基金项目(SYH-32034-0073);南京市卫生科技发展专项资金项目(YKK23209)
作者单位:1南京医科大学附属江宁医院儿科(邮编211100);2南京医科大学第二附属医院儿科;3南京医科大学第四附属医院儿科
作者简介:侯维玲(1983),女,硕士在读,主要从事儿科学方面研究。E-mail:13770698345@163.com
△通信作者 E-mail:njaiqing@njmu.edu.cn
摘要:目的 探讨锌指蛋白281(ZNF281)在高糖(HG)诱导的肾小管上皮细胞(RTECs)上皮间质转化(EMT)和细胞外基质(ECM)合成中的作用及机制。方法 使用HG诱导RTECs以构建糖尿病肾病模型,分为Control组、HG组和Mannitol组,使用CCK-8检测细胞增殖活力。使用小干扰RNA(siRNA)在HG诱导的RTECs中敲低ZNF281的表达水平,分为Control组、HG组、HG+ZNF281 siRNA组和HG+ZNF281 vector组。使用腺苷单磷酸活化蛋白激酶(AMPK)激动剂阿卡地新(AICAR)活化AMPK,分为Control组、HG组、HG+AICAR组和HG+二甲基亚砜组。实时荧光定量PCR和蛋白免疫印迹法检测ZNF281、EMT和ECM合成相关指标表达水平。结果 与Control组相比,HG组ZNF281、波形蛋白(vimentin)、α-平滑肌肌动蛋白(α-SMA)、纤维连接蛋白(FN)和Ⅰ型胶原蛋白(ColⅠ)的蛋白和mRNA表达水平升高,E-钙黏蛋白(E-cadherin)表达水平降低;与HG组相比,HG+ZNF281 siRNA组和HG+AICAR组的EMT和ECM合成相关指标的蛋白和mRNA表达水平发生明显变化,同时HG+AICAR组ZNF281的蛋白和mRNA表达水平较HG组降低;在使用AICAR和转染ZNF281过表达质粒共处理的细胞中,AICAR+ZNF281组vimentin、α-SMA、FN、ColⅠ的表达水平较空载组升高,E-cadherin表达水平降低。结论 AMPK通过负向调控ZNF281的表达水平进而抑制HG诱导的RTECs中EMT和ECM合成。
关键词:糖尿病肾病;Tristetraprolin蛋白;上皮-间质转化;细胞外基质;AMP活化蛋白激酶类;肾小管间质纤维化;锌指蛋白281
中图分类号:R587.24,R34文献标志码:ADOI:10.11958/20240031
Zinc finger protein 281 inhibits high glucose-induced epithelial-mesenchymal transition and extracellular matrix synthesis in renal tubular epithelial cells
HOU Weiling1, QIAO Yunyang2, WU Xiaoyun2, SHI Huimin2, QU Gaoting2, ZHANG Aiqing3△
1 Department of Pediatrics, the Affiliated Jiangning Hospital of Nanjing Medical University, Nanjing 211100, China;
2 Department of Pediatrics, the Second Affiliated Hospital of Nanjing Medical University; 3 Department of Pediatrics,
the Fourth Affiliated Hospital of Nanjing Medical University
△Corresponding Author E-mail: njaiqing@njmu.edu.cn
Abstract: Objective To investigate the role and mechanism of zinc finger protein 281 (ZNF281) in high glucose (HG)-induced epithelial-mesenchymal transition (EMT) and extracellular matrix (ECM) synthesis in renal tubular epithelial cells (RTECs). Methods HG induced RTECs were used to construct a diabetic kidney disease cell model, and cells were divided into the control group, the HG group and the mannitol group. Cell proliferation viability was detected by CCK-8. The expression of ZNF281 was knocked down in HG-treated RTECs using small interfering RNA (siRNA). HG-induced RTECs after knockdown of ZNF281 were divided into the control group, the HG group, the HG+ZNF281 siRNA group and the HG+ZNF281 vector group. Adenosine monophosphate-activated protein kinase (AMPK) was activated using AMPK agonist, acadexin (AICAR), and then cells were divided into the control group, the HG group, the HG+AICAR group and the HG+dimethyl sulfoxide group. The expression levels of ZNF281, EMT and ECM synthesis-related indexes were detected by qPCR and Western blot assay. Results Compared with the control group, the protein and mRNA expression levels of vimentin, α-smooth muscle actin (α-SMA), fibronectin (FN) and collagen Ⅰ (Col Ⅰ) were significantly higher, and the expression of E-cadherin was significantly lower in the HG group. Compared with the HG group, the protein and mRNA expression levels of EMT and ECM synthesis-related indexes were significantly changed in the HG+ZNF281 siRNA group and the HG+AICAR group. The protein and mRNA expression levels of ZNF281 were significantly reduced in the HG+AICAR group compared with the HG group. In cells co-treated with AICAR and transfected with ZNF281 plasmid, the expression levels of vimentin, α-SMA, FN and Col Ⅰ were significantly higher in the AICAR+ZNF281 group, and E-cadherin was significantly lower compared with that of the vector group. Conclusion AMPK inhibits EMT and ECM synthesis in HG-treated RTECs by negatively regulating the expression level of ZNF281.
Key words: diabetic nephropathies; tristetraprolin; epithelial-mesenchymal transition; extracellular matrix; AMP-activated protein kinases; tubulointerstitial fibrosis; zinc finger protein 281
糖尿病肾病(diabetic kidney disease,DKD)是糖尿病最常见的并发症之一,以大量蛋白尿和肾小球滤过率降低为特征,是终末期肾病(end-stage renal disease,ESRD)的主要原因之一,已经成为全球性的公共卫生问题[1-2]。肾脏纤维化是公认的DKD的病理路径,也是进展至ESRD的必经阶段,其病理基础包括细胞外基质(extracellular matrix,ECM)合成增多并沉积[3]。肾小管上皮细胞(renal tubular epithelial cells,RTECs)作为肾脏纤维化主要的效应细胞之一,在DKD发病早期可异常活化和增殖,促进肾小管间质纤维化(tubular interstitial fibrosis,TIF)及ECM合成,是DKD病理改变的关键环节[4],而上皮间质转化(epithelial-mesenchymal transition,EMT)是TIF发生和发展的重要机制[5],确定DKD的发病机制并找寻新的治疗靶点是目前亟待解决的问题[6]。EMT和ECM合成相关蛋白包括E-钙黏蛋白(E-cadherin)、波形蛋白(vimentin)、α-平滑肌肌动蛋白(α-SMA)、纤维连接蛋白(fibronectin,FN)和Ⅰ型胶原蛋白(collagenⅠ,ColⅠ)。锌指蛋白(zinc finger protein,ZNF)是一类转录因子,能与多种基因启动子中的GC区域结合,在基因调控中发挥关键作用[7-8]。ZNF是组织发育的关键调控因子,在肿瘤相关疾病中更多是以高水平表达[9-10]。ZNF281可能与EMT相关,在炎症性肠病中沉默ZNF281可明显减少炎性因子释放,降低纤维化相关基因的表达水平,并缓解由炎症引起的组织细胞形态变化[11]。生物信息学分析发现,在糖尿病和DKD患者血清测序表达谱中腺苷单磷酸活化蛋白激酶(AMPK)明显富集[12],通过激活AMPK可以保护细胞免受炎症、凋亡、各类应激及EMT的影响[13]。AMPK可进一步磷酸化多种下游转录因子,参与调控糖代谢、脂代谢及细胞增殖等过程[14-15]。本研究从AMPK出发,通过高糖(high glucose,HG)诱导RTECs建立DKD细胞模型,探讨ZNF281在DKD中的调控机制,为DKD的临床诊治提供新策略。
1 材料与方法
1.1 材料
1.1.1 细胞系 小鼠RTECs由东南大学附属中大医院肾脏病中心馈赠。
1.1.2 主要试剂和仪器 细胞培养所用培养基、胰酶、胎牛血清和磷酸盐缓冲液均购自美国Gibco公司;AMPK激动剂阿卡地新(AICAR)购自MCE;细胞总蛋白提取及样本制备所需的裂解液和蛋白上样缓冲液购自上海碧云天生物技术有限公司;蛋白免疫印迹法(Western blot)所使用的凝胶、电泳缓冲液、快速转膜液及电泳设备均购自赛维尔生物科技有限公司,凝胶成像仪购自Bio-Rad公司,聚偏氟乙烯(PVDF)膜购自美国密理博公司,ZNF281抗体购自武汉bioswamp公司,FN、ColⅠ、AMPK及磷酸化AMPK(p-AMPK)抗体购自英国Abcam公司,E-cadherin、vimentin、α-SMA、GAPDH抗体以及辣根过氧化物酶标记的山羊抗兔IgG和山羊抗鼠IgG二抗均购自Affinity公司;TRIzol试剂盒购自美国Invitrogen公司,逆转录实时荧光定量PCR(qPCR)相关试剂盒、CCK-8细胞增殖检测试剂盒均购自南京诺唯赞生物科技股份有限公司,PCR仪为Roche LightCycler 480;ZNF281小干扰RNA(siRNA)及相应的空载对照(vector)由广州锐博生物技术有限公司构建,同时配合使用该公司提供的细胞转染试剂盒;ZNF281过表达质粒由南京科瑞斯生物科技有限公司构建。
1.2 方法
1.2.1 CCK-8检测细胞增殖 为筛选构建DKD细胞模型的最适干预方案,使用5.5、30、50、80 mmol/L葡萄糖干预细胞并设置12、24、36、48 h 4个时间点。取对数生长期的RTECs并接种于96孔板中,预留空白对照孔,每孔的终体积为100 μL,当干预达到实验需要后每孔加入10 μL CCK-8溶液并置于细胞培养箱中孵育4 h,使用酶标仪测定各孔450 nm处光密度(OD450)值,检测细胞增殖情况。
1.2.2 细胞处理及分组 RTECs培养于37 ℃、含5%CO2的细胞培养箱中,在细胞密度达到80%时接种于6孔板或12孔板中,并在适当时间依实验需要处理细胞。对于DKD细胞模型的建立,将细胞分为3组:(1)对照(Control)组。使用低浓度(5.5 mmol/L)葡萄糖培养基培养。(2)HG组。使用葡萄糖配置HG培养基(30 mmol/L)干预细胞作为DKD模型。(3)甘露醇(Mannitol)组。使用5.5 mmol/L葡萄糖和24.5 mmol/L甘露醇配置与HG组相同渗透压的培养基,以排除HG的高渗透压对细胞的影响。ZNF281的转染实验中将RTECs分为以下4组:(1)Control组为5.5 mmol/L葡萄糖培养细胞。(2)HG组为30 mmol/L葡萄糖培养细胞。(3)HG+ZNF281 siRNA组为在HG诱导的RTECs中转染ZNF281 siRNA。(4)HG+ZNF281 vector组为在HG诱导的RTECs中转染ZNF281 siRNA相应的阴性对照。AICAR对细胞的处理实验中将RTECs分为以下4组:(1)Control组为5.5 mmol/L葡萄糖培养。(2)HG组为30 mmol/L葡萄糖培养细胞。(3)HG+AICAR组使用AICAR处理HG干预的RTECs。(4)HG+二甲基亚砜(DMSO)组在HG诱导的细胞中加入与配置AICAR所需的DMSO相应浓度和剂量以排除AICAR配置液对细胞的影响。对于AICAR和转染ZNF281过表达质粒共处理实验中将RTECs分为2组:(1)AICAR+ZNF281 vector组为使用AICAR处理细胞并共转染ZNF281过表达质粒的相应空载。(2)AICAR+ZNF281组为使用AICAR处理细胞并转染ZNF281过表达质粒。在干预24 h后收集细胞并用于后续实验。
1.2.3 细胞转染 在6孔板或12孔板中,当细胞生长的密度达到约70%时进行细胞转染,使用锐博生物提供的细胞转染试剂盒并依据说明书进行操作。使用缓冲液及siRNA、阴性参照或质粒配制终浓度为20 nmol/L的转染混合物,期间需轻柔震荡并混匀,室温孵育15 min后,依据实验分组进行转染,通过qPCR和Western blot验证转染效率。用于转染的ZNF281 siRNA的序列:5′-GTACTCTGGCAAATCAGAA-3′,其阴性参照序列为广州锐博生物技术有限公司专利。
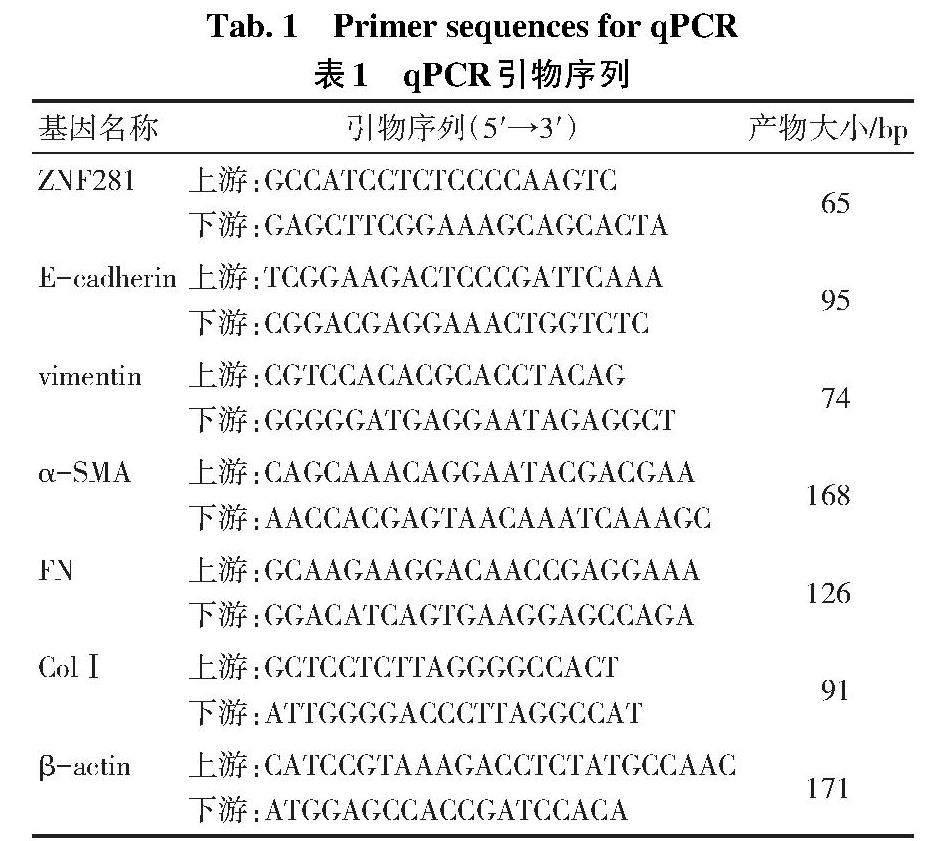
1.2.4 qPCR检测ZNF281、EMT和ECM合成相关基因表达水平 使用TRIzol裂解细胞并提取总RNA,测定RNA浓度后,依据逆转录检测试剂盒说明书将总RNA定量并逆转录为cDNA,使用ChamQ SYBR qPCR Master Mix试剂盒进行后续的扩增,以β-actin作为内参。PCR反应程序为:95 ℃ 30 s预变性;95 ℃ 10 s,69 ℃ 30 s,循环40次。95 ℃ 15 s,60 ℃ 1 min,95 ℃ 15 s以构建熔解曲线。采用2-ΔΔCt法计算各组细胞目的基因的相对表达水平。引物均由上海捷瑞生物工程有限公司构建并进行了验证,引物序列见表1。[Tab.1 Primer sequences for qPCR
1.2.5 Western blot检测ZNF281、AMPK、EMT和ECM相关蛋白表达 细胞总蛋白的提取全程于冰上进行,使用加入蛋白酶抑制剂的裂解缓冲液裂解细胞,刮下细胞并收集细胞悬浮液,随后在4 ℃下以12 500×g高速离心20 min,收集上清液,测定蛋白浓度。制备好蛋白样品并保存于-80 ℃冰箱内或加载至预制好的凝胶中,调节电压为160 V进行电泳分离蛋白,约40 min后,使用快速免冰浴转膜液将凝胶上的蛋白在300 mA恒定电流下转移到PVDF膜上,使用脱脂奶粉配置封闭液并在室温下封闭1 h,封闭结束后洗膜并加入配置好的一抗(配置比例均为1∶1 000),在4 ℃冰箱中孵育过夜。次日,复温并洗膜后加入相应二抗(配置比例为1∶10 000),于室温下孵育1 h,再次洗膜,加入增强型化学发光液并置于Bio-Rad凝胶成像仪下,使用Image Lab 4.0曝光并记录蛋白条带。使用ImageJ 1.53t分析蛋白条带的灰度值,以GAPDH作为内参,将对照组归一化后计算目的蛋白的相对表达量,每组实验重复至少3次并合并分析蛋白表达量。
1.3 统计学方法 使用GraphPad Prism 9.3软件进行数据分析,计量资料以均数±标准差([x] ±s)表示,2组间比较采用t检验,多组间比较使用单因素方差分析,组间多重比较采用Sidak法,P<0.05为差异有统计学意义。
2 结果
2.1 ZNF281在HG诱导的RTECs中高表达 CCK-8结果显示,当葡萄糖浓度为30 mmol/L时,干预24 h的RTECs增殖活力高于其他干预方案,见图1。HG组RTECs中ZNF281、vimentin、α-SMA、FN、ColⅠ的mRNA和蛋白表达量较Control组升高,E-cadherin表达水平降低(均P<0.05),Mannitol组相关指标表达水平较Control组无明显变化,见图2、表2。
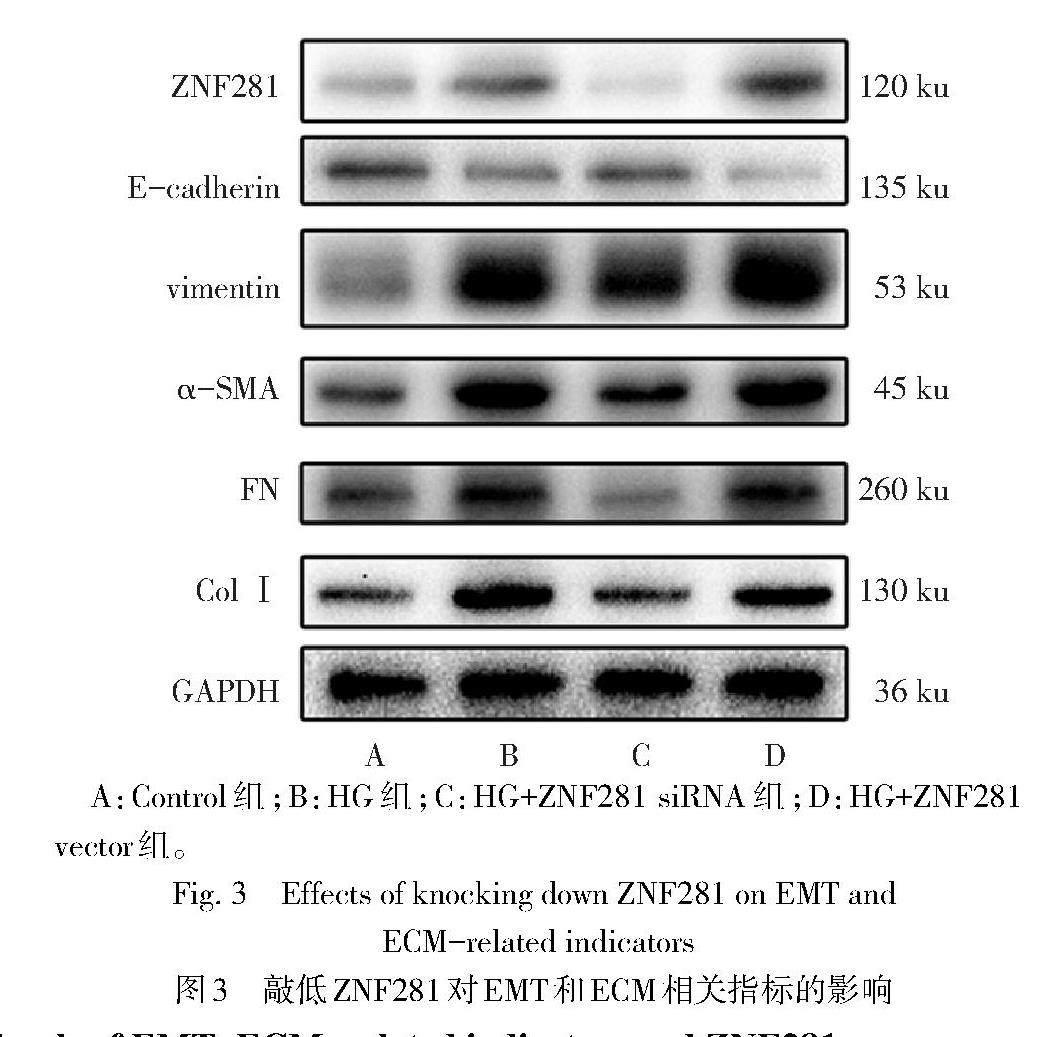
2.2 敲低ZNF281可抑制HG诱导RTECs的EMT和ECM合成 与HG组相比,HG+ZNF281 siRNA组细胞中ZNF281表达水平降低,vimentin、α-SMA、FN、ColⅠ的mRNA和蛋白表达水平降低,E-cadherin表达水平升高(均P<0.05),HG+ZNF281 vector组相关指标表达水平与HG组比较均无统计学意义,见图3、表3。CCK-8结果显示,Control组、HG组、HG+ZNF281 siRNA组、HG+ZNF281 vector组OD450分别为0.64±0.06、1.78±0.03、1.37±0.07、1.85±0.09(F=269.400,P<0.01),与HG组比较,HG+ZNF281 siRNA组细胞增殖活力降低(P<0.05)。
2.3 激活AMPK可抑制HG诱导RTECs的EMT和ECM合成 Control组、HG组、Mannitol组p-AMPK/AMPK蛋白表达水平分别为1.00±0.05、0.66±0.13、1.02±0.14(F=9.025,P<0.05);与Control组比较,HG组p-AMPK/AMPK明显下调(P<0.05),AMPK失活,见图4。进一步在HG诱导的RTECs中使用AMPK激动剂AICAR干预细胞,CCK-8结果显示,Control组、HG组、HG+AICAR组、HG+DMSO组细胞OD450分别为0.57±0.04、1.64±0.08、1.22±0.12、1.66±0.19,其差异有统计学意义(F=71.470,P<0.05);与HG组相比,HG+AICAR组细胞增殖活力明显降低(P<0.05);与HG组相比,HG+AICAR组p-AMPK/AMPK蛋白表达水平升高(P<0.05),AMPK活化;与HG组相比,HG+AICAR组vimentin、α-SMA、FN、ColⅠ的mRNA和蛋白表达水平降低(P<0.05),E-cadherin表达水平升高(P<0.05),HG+DMSO组相关指标较HG组差异无统计学意义,见表4、图5。
2.4 AMPK通过调控ZNF281抑制HG诱导RTECs的EMT和ECM合成 HG+AICAR组ZNF281的mRNA和蛋白表达水平较HG组降低(P<0.05),见图6、表4。进一步在AICAR处理的RTECs中共转染ZNF281,结果显示,共处理组ZNF281表达水平较AICAR+ZNF281 vector升高,vimentin、α-SMA、FN、ColⅠ的mRNA和蛋白表达水平升高,E-cadherin表达水平降低(均P<0.05),见表5、图7;
CCK-8结果显示,AICAR+ZNF281 vector细胞OD450为1.34±0.07,而共处理组的OD450为2.19±0.12,细胞增殖活力显著升高(t=12.050,P<0.05)。
3 讨论
目前DKD在全球糖尿病患者中的发病率逐年上升[2]。TIF是各类慢性肾脏病进展到ESRD的共同阶段,早期并有效地减轻炎症反应,抑制EMT和减少ECM过度沉积,对于改善患者预后具有重要意义。有关ZNF281在肾脏病理中作用的研究报道尚少,其与TIF的发生发展关系不明确,本研究评估并确定了ZNF281在DKD模型中的表达水平及其敲低对DKD的影响,并初步探讨了调控机制,结果发现ZNF281表达受AMPK控制,AMPK激活导致ZNF281表达下调,抑制了HG诱导的RTECs中EMT的发生和ECM过度沉积,并与细胞增殖活力降低相关。
ZNF281属于Krüppel样锌指转录因子家族,具有转录因子活性,因含有指状结构域得名,广泛分布于全身多个组织[16]。研究表明其与胚胎干细胞多能性、DNA损伤修复有关[9,17]。在DKD病理基础中,上皮细胞获得间充质特征,运动性和侵袭性明显增强,加速DKD进展。目前为止转录因子Snail、Slug等被广泛认为是EMT调节的核心因子。研究证实原发性结直肠癌样本中ZNF281的表达明显上调,其失活可明显抑制原癌基因诱导的EMT[18]。在小鼠纤维化结肠和经转化生长因子-β1诱导的人源性结肠成纤维细胞中,ZNF281的表达明显增加,且与成纤维细胞活化、分化相关途径的基因表达存在关联[19]。在炎症性肠病中,ZNF281的表达与疾病严重程度呈正相关,沉默ZNF281可以明显降低纤维化相关蛋白的表达,并抑制炎症因子释放,缓解由炎症引起的形态学改变[11];而肾小管间质炎症是DKD病理改变的关键环节[20],因此笔者推测ZNF281可能是EMT调控网络的一个重要组成部分,在慢性肾脏病中同样发挥重要的调节作用,具有成为DKD临床诊治新靶点的潜力。本研究结果显示ZNF281在HG诱导的RTECs中表达水平明显上调,进一步探究ZNF281在HG诱导的RTECs中EMT和ECM合成的作用,在HG诱导的RTECs中敲低ZNF281可以明显抑制EMT和ECM的合成,并与细胞增殖活力的降低密切相关。本研究为ZNF281在肾脏病领域作为EMT相关分子提供了新的证据。
高浓度的葡萄糖具有内在毒性,可引发肾小管间质纤维化和肾小球系膜细胞增殖[21-22]。本研究使用HG诱导RTECs,通过CCK-8检测细胞增殖,确定糖浓度为30 mmol/L并干预24 h作为DKD细胞模型诱导方案,成功建立了DKD细胞模型。AMPK作为代谢应激中高度保守的能量平衡调节因子,在溶酶体表面感知葡萄糖,并进一步磷酸化下游转录因子,参与调控糖代谢、脂代谢及细胞增殖等过程[23]。AICAR是一种腺苷类似物,作为AMPK激动剂可以调节葡萄糖和脂质代谢,保护细胞免受活性氧刺激和EMT的影响,并抑制促炎细胞因子的产生[24]。本研究结果显示,在HG诱导的RTECs中,AICAR和ZNF281 siRNA的处理对于DKD细胞模型中的EMT和ECM合成存在类似的效应。AMPK可以通过磷酸化下游转录因子发挥效应,因此笔者提出假设:AMPK通过调控ZNF281的表达进而抑制EMT和ECM合成。在HG诱导的RTECs中使用AICAR处理后,ZNF281的表达水平明显下调,确定AMPK对ZNF281的调控关系;在使用ZNF281过表达质粒共处理后,AICAR对DKD细胞模型中EMT和ECM沉积的治疗作用发生了逆转,进一步表明AMPK通过ZNF281调节EMT的发生和ECM合成。
综上,本研究提供了ZNF281作为肾脏纤维化诱导因子的证据,确定AMPK与ZNF281的相互调控关系,AMPK通过负向调控ZNF281的表达来抑制HG诱导的RTECs中EMT的发生和ECM合成。上述结论加深了对TIF发展机制的理解,并为DKD临床诊治提供了理论依据。然而本研究仍存在一定不足,如ZNF281在DKD动物模型中介导的效应,以及ZNF281的下游作用靶点尚不清楚,期望能够在后续的研究中有更多的发现。
参考文献
[1] THURLOW J S,JOSHI M,YAN G,et al. Global epidemiology of end-stage kidney disease and disparities in kidney replacement therapy[J]. Am J Nephrol,2021,52(2):98-107. doi:10.1159/000514550.
[2] ALICIC R Z,ROONEY M T,TUTTLE K R. Diabetic kidney disease:challenges,progress,and possibilities[J]. Clin J Am Soc Nephrol,2017,12(12):2032-2045. doi:10.2215/CJN.11491116.
[3] HUMPHREYS B D. Mechanisms of renal fibrosis[J]. Annu Rev Physiol,2018,80:309-326. doi:10.1146/annurev-physiol-022516-034227.
[4] JIANG S,SU H. Cellular crosstalk of mesangial cells and tubular epithelial cells in diabetic kidney disease[J]. Cell Commun Signal,2023,21(1):288. doi:10.1186/s12964-023-01323-w.
[5] LOVISA S,ZEISBERG M,KALLURI R. Partial epithelial-to-mesenchymal transition and other new mechanisms of kidney fibrosis[J]. Trends Endocrinol Metab,2016,27(10):681-695. doi:10.1016/j.tem.2016.06.004.
[6] TUTTLE K R,AGARWAL R,ALPERS C E,et al. Molecular mechanisms and therapeutic targets for diabetic kidney disease[J]. Kidney Int,2022,102(2):248-260. doi:10.1016/j.kint.2022.05.012.
[7] MAYER D,STADLER M B,RITTIRSCH M,et al. Zfp281 orchestrates interconversion of pluripotent states by engaging Ehmt1 and Zic2[J]. EMBO J,2020,39(2):e102591. doi:10.15252/embj.2019102591.
[8] LU C,GE T,SHAO Y,et al. ZNF281 drives hepatocyte senescence in alcoholic liver disease by reducing HK2-stabilized PINK1/Parkin-mediated mitophagy[J]. Cell Prolif,2023,56(3):e13378. doi:10.1111/cpr.13378.
[9] ISHIUCHI T,OHISHI H,SATO T,et al. Zfp281 shapes the transcriptome of trophoblast stem cells and is essential for placental development[J]. Cell Rep,2019,27(6):1742-1754.e6. doi:10.1016/j.celrep.2019.04.028.
[10] DENG Y,PENG D,XIAO J,et al. Inhibition of the transcription factor ZNF281 by SUFU to suppress tumor cell migration[J]. Cell Death Differ,2023,30(3):702-715. doi:10.1038/s41418-022-01073-1.
[11] PIERDOMENICO M,PALONE F,CESI V,et al. Transcription factor ZNF281:a novel player in intestinal inflammation and fibrosis[J]. Front Immunol,2018,9:2907. doi:10.3389/fimmu.2018.02907.
[12] HUANG C,DING L,JI J,et al. Expression profiles and potential roles of serum tRNA-derived fragments in diabetic nephropathy[J]. Exp Ther Med,2023,26(1):311. doi:10.3892/etm.2023.12010.
[13] HAN Y C,TANG S Q,LIU Y T,et al. AMPK agonist alleviate renal tubulointerstitial fibrosis via activating mitophagy in high fat and streptozotocin induced diabetic mice[J]. Cell Death Dis,2021,12(10):925. doi:10.1038/s41419-021-04184-8.
[14] PAQUETTE M,EL-HOUJEIRI L,C ZIRDEN L,et al. AMPK-dependent phosphorylation is required for transcriptional activation of TFEB and TFE3[J]. Autophagy,2021,17(12):3957-3975. doi:10.1080/15548627.2021.1898748.
[15] WANG Y,LUO M,WANG F,et al. AMPK induces degradation of the transcriptional repressor PROX1 impairing branched amino acid metabolism and tumourigenesis[J]. Nat Commun,2022,13(1):7215. doi:10.1038/s41467-022-34747-y.
[16] WOO A J,PATRY C A,GHAMARI A,et al. Zfp281(ZBP-99) plays a functionally redundant role with Zfp148(ZBP-89)during erythroid development[J]. Blood Adv,2019,3(16):2499-2511. doi:10.1182/bloodadvances.2018030551.
[17] WANG Y,MA B,LIU X,et al. ZFP281-BRCA2 prevents R-loop accumulation during DNA replication[J]. Nat Commun,2022,13(1):3493. doi:10.1038/s41467-022-31211-9.
[18] HAHN S,JACKSTADT R,SIEMENS H,et al. SNAIL and miR-34a feed-forward regulation of ZNF281/ZBP99 promotes epithelial-mesenchymal transition[J]. EMBO J,2013,32(23):3079-3095. doi:10.1038/emboj.2013.236.
[19] LAUDADIO I,BASTIANELLI A,FULCI V,et al. ZNF281 promotes colon fibroblast activation in TGFβ1-induced gut fibrosis[J]. Int J Mol Sci,2022,23(18):10261. doi:10.3390/ijms231810261.
[20] YANG W X,LIU Y,ZHANG S M,et al. Epac activation ameliorates tubulointerstitial inflammation in diabetic nephropathy[J]. Acta Pharmacol Sin,2022,43(3):659-671. doi:10.1038/s41401-021-00689-2.
[21] ZENI L,NORDEN A,CANCARINI G,et al. A more tubulocentric view of diabetic kidney disease[J]. J Nephrol,2017,30(6):701-717. doi:10.1007/s40620-017-0423-9.
[22] 吴玮熙,刘帅辉,周赛君,等. 高糖通过miR-192调控CAV-1/EGF影响肾小球系膜细胞增殖、迁移及细胞外基质形成的机制研究[J]. 天津医药,2022,50(10):1020-1025. WU W X,LIU S H,ZHOU S J,et al. Effects of high glucose on the proliferation and migration of mesangial cells and extracellular matrix formation by regulating the CAV-1/EGF through miR-192[J].Tianjin Med J,2022,50(10):1020-1025. doi:10.11958/20220268.
[23] LIN S C,HARDIE D G. AMPK: sensing glucose as well as cellular energy status[J]. Cell Metab,2018,27(2):299-313. doi:10.1016/j.cmet.2017.10.009.
[24] TUKHOVSKAYA E A,SHAYKHUTDINOVA E R,PAKHOMOVA I A,et al. AICAR improves outcomes of metabolic syndrome and type 2 diabetes induced by high-fat diet in C57Bl/6 male mice[J]. Int J Mol Sci,2022,23(24):15719. doi:10.3390/ijms232415719.
(2024-01-04收稿 2024-02-26修回)
(本文编辑 李国琪)
猜你喜欢
传染病信息(2022年3期)2022-07-15
肝博士(2021年1期)2021-03-29
中国甜菜糖业(2020年1期)2020-12-07
学苑创造·A版(2020年12期)2020-01-07
中国外汇(2019年15期)2019-10-14
江苏卫生保健(2018年7期)2018-07-31
中国乳业(2016年4期)2016-11-07
作文教学研究(2016年1期)2016-07-05
肿瘤影像学(2015年3期)2015-12-09
医学研究杂志(2015年6期)2015-07-01
