
基于高温半致死温度的13个牡丹品种耐热性评价
2024-07-04 12:59:05余昕陈法志刘忠陈志伟戢小梅李秀丽
湖北农业科学 2024年6期
余昕 陈法志 刘忠 陈志伟 戢小梅 李秀丽
摘要:以13个代表性牡丹(Paeonia suffruticosa Andr.)品种为试材,利用电导率法、Logistic方程研究高温半致死温度(LT50),以期评价牡丹种质资源的耐热性。结果表明,在逐步高温处理过程中,13个牡丹品种的细胞伤害率呈典型的S形曲线增长趋势,经显著性检验符合Logistic方程。13个牡丹品种的LT50为51.31~59.73 ℃,其中B24的LT50最高,为59.73 ℃,而海黄的LT50最低,为51.31 ℃。利用有序样品最优分割聚类法,将13个牡丹品种分为3个类群,B24、太平红、B10、凤丹荷和凤丹白属于耐热类型,LT50平均值为57.70 ℃;香玉、莲鹤、五星玉、岛锦和垫江红属于中度耐热类型,LT50平均值为56.32 ℃;ZB1、泼墨紫和海黄属于不耐热类型,LT50平均值为53.32 ℃。
关键词:牡丹(Paeonia suffruticosa Andr.); 细胞伤害率; Logistic方程; 高温半致死温度; 耐热性
中图分类号:S685.11 文献标识码:A
文章编号:0439-8114(2024)06-0116-05
DOI:10.14088/j.cnki.issn0439-8114.2024.06.018 开放科学(资源服务)标识码(OSID):
Evaluation of heat resistance of 13 tree peony varieties based on semi-lethal high temperature
YU Xin, CHEN Fa-zhi, LIU Zhong, CHEN Zhi-wei, JI Xiao-mei, LI Xiu-li
(Institute of Forestry and Fruit Tree, Wuhan Academy of Agricultural Sciences, Wuhan 430075, China)
Abstract: In order to evaluate the heat resistance of tree peony(Paeonia suffruticosa Andr.) germplasm resources, 13 representative tree peony cultivars were selected as experimental materials, and the semi-lethal high temperature (LT50) was studied by the conductivity method and Logistic equation. The results showed that with the treating temperatures higher step-by-step, the cell damage rate of 13 tree peony cultivars showed a typical S curve increasing trend, and the significance test conformed to the Logistic equation. The LT50 of 13 tree peony cultivars ranged from 51.31 ℃ to 59.73 ℃, among which B24 had the highest LT50 of 59.73 ℃, while Haihuang had the lowest LT50 of 51.31 ℃. Based on sequential clustering method, 13 tree peony cultivars could be divided into 3 groups: B24, Taipinghong, B10, Fengdanhe and Fengdanbai belonged to the heat resistant type, with an average LT50 of 57.70 ℃; Xiangyu, Lianhe, Wuxingyu, Daojin and Dianjianghong belonged to the moderate heat resistant type, with an average LT50 of 56.32 ℃; ZB1, Pomozi and Haihuang belonged to the heat sensitive type, with an average LT50 of 53.32 ℃.
Key words: tree peony(Paeonia suffruticosa Andr.); cell damage rate; Logistic equation; semi-lethal high temperature; heat resistance
牡丹(Paeonia suffruticosa Andr.)属于芍药科(Paeoniaceae)芍药属(Paeonia)牡丹组(Paeonia sect. Moutan DC.)植物,被称为花中之王,是世界著名花卉,也是中国十大名花之一,具有较高的观赏价值[1]。中国不仅是牡丹野生种的惟一产地,也是栽培品种的起源和演化中心,已有数千年的自然生长和1 500多年的人工栽培历史[2]。牡丹适应性强,耐干旱,能够种植在非耕性的荒山荒坡上,在水土保持、防风固沙、改善土壤品质等方面具有重要作用[3]。
牡丹原生地位于中国境内的河南、西藏和湖北等地的高海拔地区,性喜冷凉干燥的环境,在年平均气温12~15 ℃的黄河中下游地区生长良好,26 ℃以上则停止生长,进入暂时休眠[4]。中国多地35 ℃以上高温主要集中在6月中旬到8月中旬,随着高温时间的延长,叶片出现失绿、黄化、焦枯、提早落叶,直至植株死亡[5,6]。随着全球气候变暖,极端高温事件频繁发生,北部中纬度地区更容易受到影响,有些地区打破了长期气温记录的5 ℃之高[7]。此外,夏季日数、热夜日数、暖夜日数和暖昼日数也均呈显著上升趋势[8],高温已成为影响牡丹生长的重要环境限制因子,不仅使其在原适宜生长的地区面临挑战,也严重抑制着牡丹在长江中下游地区的推广应用。
研究人员从抗性育种[9]、抗性生理[10]、基因调控[11]、转录组[12]和蛋白质组[13]等方面开展牡丹耐热性研究,发现高温不仅影响牡丹表型变化,而且还会引起体内细胞稳态失衡、影响物质代谢和生物合成等过程,抑制植物的生长和发育,严重影响了牡丹的生物学特性及观赏性。植物细胞膜具有选择透过性,对维持细胞结构稳定和调节生理代谢活动起着至关重要的作用。当植物受到高温胁迫时,细胞膜出现分子内化学键松动、蛋白质变性或脂肪酸组分发生改变等现象,导致膜渗透性增大,细胞内的电解质外渗不断增加[14]。细胞膜的热稳定性是评价植物耐热性的重要指标,尤其是利用电导率法和Logistic方程测定植物的高温半致死温度(LT50),已广泛用于植物的耐热性评价,比如绣球花[15]、草莓[16]、小白菜[17]、蓝莓[18]、三棱栎[19]、藤本植物[20]等均有报道。然而有关牡丹种质资源高温半致死温度的研究仍有待深入探讨。
本研究以13个牡丹品种为试材,包括从湖北省保康县、河南省洛阳市、安徽省铜陵市、四川省彭州市、湖南省邵阳市、山东省菏泽市、重庆市垫江县等地引进的栽培品种,以及资源圃中性状稳定的变异品种,具有一定的主栽品种代表性。应用电导率法和Logistic方程拟合计算,明确不同种质资源的高温半致死温度,并结合有序样品最优分割聚类法对不同牡丹品种的耐热性强弱进行聚类分析,为其在园林绿化及美丽乡村建设中的应用提供参考依据。
1 材料与方法
1.1 材料
以泼墨紫(Pomozi)、岛锦(Daojin)、莲鹤(Lianhe)、凤丹荷(Fengdanhe)、五星玉(Wuxingyu)、海黄(Haihuang)、B10、香玉(Xiangyu)、B24、垫江红(Dianjianghong)、ZB1、太平红(Taipinghong)、凤丹白(Fengdanbai)共13个牡丹品种为供试植物。上述供试植物栽培于武汉市农业科学院林业果树研究所牡丹资源圃中,常规水肥管理。选取长势基本一致的6年生嫁接苗用于试验。
1.2 方法
1.2.1 细胞伤害率的测定 分别选取枝条自下而上的第2至第3个复叶顶部向下第2至第3片功能小叶,用清水和去离子水清洗干净后擦干水分,去除叶片的主叶脉和边缘,用剪刀剪取直径约为6 mm的小圆片,取0.1 g叶片样品放入10 mL的离心管中,加入9 mL去离子水,用针筒抽真空使叶片沉入离心管底部。以室温下各品种牡丹叶片的电导率作为对照(RCK),并分别在30、35、40、45、50、55、60、65 ℃的梯度水浴中恒温水浴20 min,待水浴完成后冷却至室温,采用雷磁电导仪(DDS-307A)测定不同水浴温度下的电导率(Rt);再将牡丹叶片置于沸水中煮沸15 min,待水浴完成后,静置冷却至室温,测出最终电导率(R0),每组处理重复3次。按如下公式计算细胞伤害率,细胞伤害率=(Rt-RCK)/(R0-RCK)×100%[20,21]。
1.2.2 Logistic方程和高温半致死温度的确定 利用Logistic方程Y=k/(1+ae-bt),对各个处理温度与所测细胞伤害率进行拟合。其中,Y为细胞伤害率;k为细胞伤害率的饱和容量;t为处理温度,由于细胞伤害率去除了本底干扰,因此在本试验中k为100%;a、b为方程参数。为了确定a和b的值,将Logistic方程进行线性化处理,ln[(k-Y)/Y] = lna-bt,令y = ln[(k-Y)/Y],则转化为细胞伤害率(y)与处理温度(t)的直线方程。通过DPS 9.01软件回归分析的方法求得参数a、b的值以及相关系数,计算高温半致死温度,对Logistic方程进行二阶求导,并令其等于0,获得曲线的拐点t=(lna)/b,求出的t即为高温半致死温度(LT50)[20,22]。
1.3 数据分析
使用Excel 2016软件整理试验数据并绘制图表,采用SPSS 20.0软件和DPS 9.01统计软件分析数据。
2 结果与分析
2.1 不同牡丹品种细胞伤害率与处理温度的关系
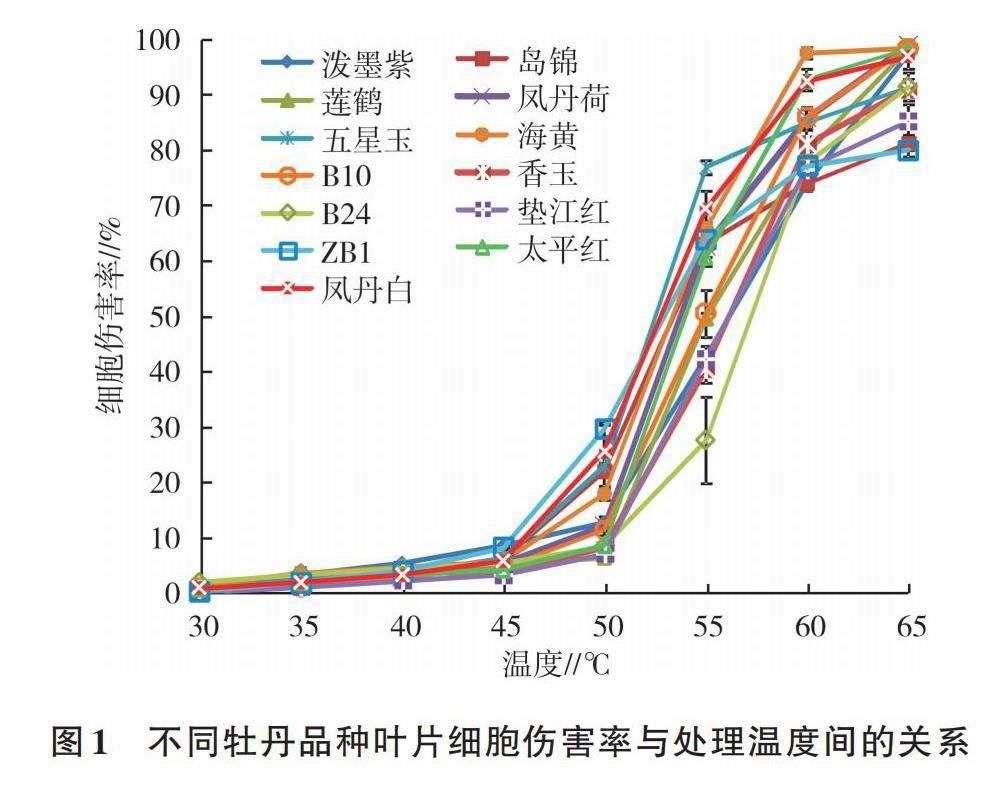
由图1可以看出,当温度在30~65 ℃时,随着处理温度的升高,13个牡丹品种叶片的细胞伤害率不断增加,呈缓慢上升-急剧上升-缓慢上升的趋势,即典型的S形曲线,不同品种间的上升快慢存在差异。在30~45 ℃,13个牡丹品种的叶片细胞伤害率增加缓慢,除ZB1增加了25.52倍外,其他品种变化较小,表明此温度区间内,牡丹叶片能自主调节并且维持细胞相对稳定的状态,但ZB1的自主调节能力较差。在45~50 ℃,ZB1、凤丹白、五星玉、岛锦、海黄的细胞伤害率上升较快,说明该温度范围对这些品种细胞稳定性的影响较大,尤其对ZB1,其细胞伤害率从8.22%上升至29.56%。在50~60 ℃,13个品种的细胞伤害率均呈急剧上升趋势,泼墨紫、岛锦、莲鹤、B24、垫江红、ZB1的细胞伤害率为70%~80%,凤丹荷、五星玉、B10、香玉的细胞伤害率为80%~90%,而海黄、太平红、凤丹白的细胞伤害率均在90%以上,其中海黄的细胞伤害率最大,达97.63%,表明牡丹叶片细胞质膜结构受损严重,细胞内电解质已出现大量外渗。当温度上升至65 ℃时,13个品种的细胞伤害率均达80%以上,其中,泼墨紫的细胞伤害率从73.85%上升至97.31%,增幅达23.46个百分点,其他品种细胞伤害率增长趋于平缓,表明此时牡丹细胞质膜遭到严重破坏,丧失了调节和选择物质进出的主动运输功能,细胞内外电解质浓度已趋于平衡。综上可知,ZB1、凤丹白、五星玉、岛锦、海黄叶片膜系统不可逆伤害的临界温度为45~60 ℃,莲鹤、泼墨紫、B24、垫江红、太平红、B10、香玉、凤丹荷叶片膜系统的临界温度为50~60 ℃。
2.2 不同牡丹品种高温半致死温度的确定
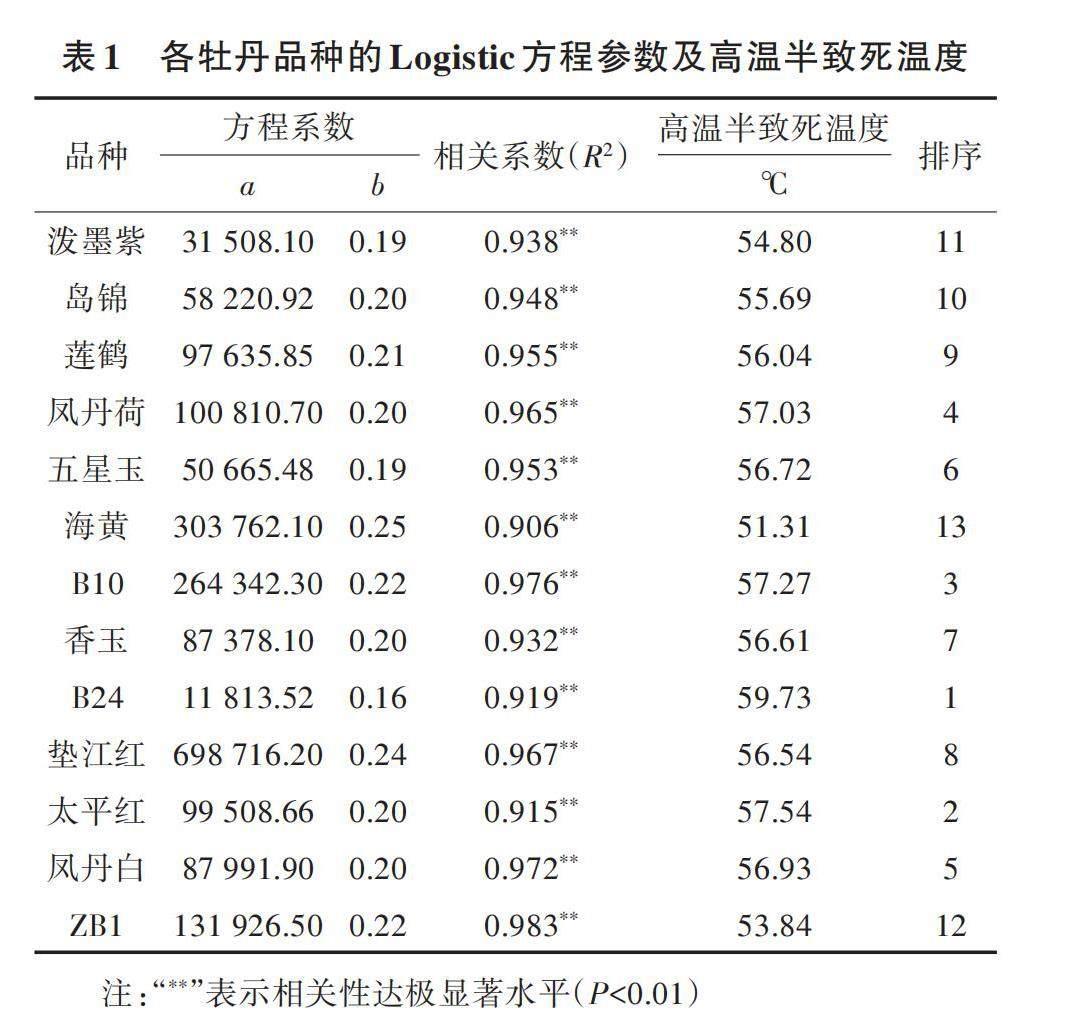
对不同温度处理下13个牡丹品种的细胞伤害率进行一元回归分析和Logistic方程拟合,确定了Logistic方程参数和相关系数,并计算高温半致死温度,结果如表1所示。13个牡丹品种的相关系数(R2)为0.906~0.983,达极显著水平,方程拟合程度较好。13个牡丹品种的LT50为51.31~59.73 ℃,平均LT50为56.16 ℃。由于高温半致死温度与牡丹耐热性呈正相关,可见13个牡丹品种的耐热性表现为B24(59.73 ℃)>太平红(57.54 ℃)>B10(57.27 ℃)>凤丹荷(57.03 ℃)>凤丹白(56.93 ℃)>五星玉(56.72 ℃)>香玉(56.61 ℃)>垫江红(56.54 ℃)>莲鹤(56.04 ℃)>岛锦(55.69 ℃)>泼墨紫(54.80 ℃)>ZB1(53.84 ℃)>海黄(51.31 ℃)。
2.3 不同牡丹品种的耐热性聚类分析
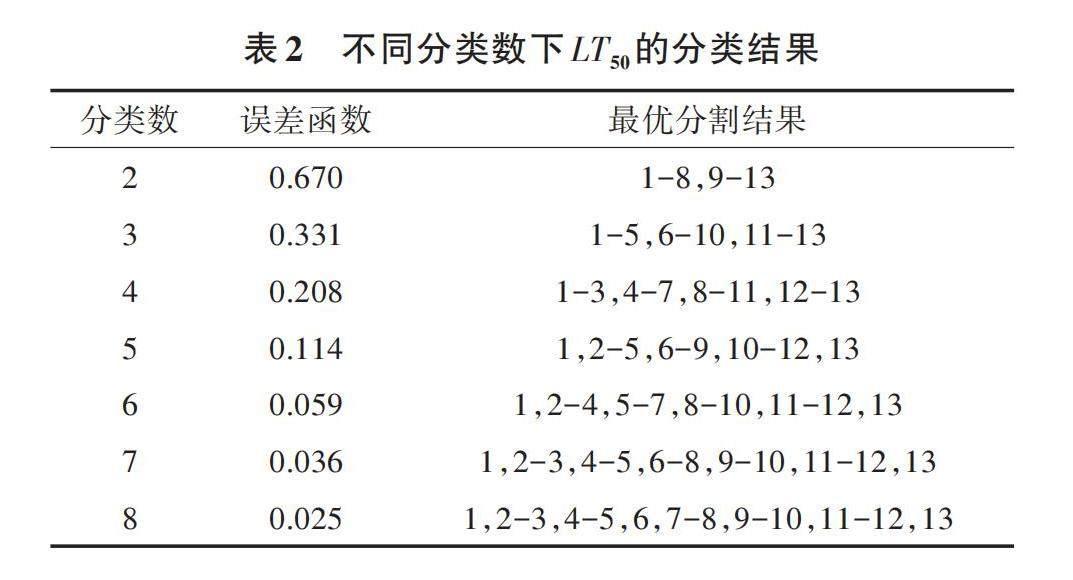
进一步通过有序样品最优分割聚类法对13个牡丹品种的耐热性强弱进行分类,结果见表2、图2和表3。分类数与误差函数曲线的拐点在3~4,将13个牡丹品种的耐热性按LT50的排序分为3类,排序1~5的B24、太平红、B10、凤丹荷、凤丹白为耐热品种(R),排序6~10的五星玉、香玉、垫江红、莲鹤、岛锦为中度耐热品种(M),排序11~13的泼墨紫、ZB1、海黄为不耐热品种(S)。
3 讨论
在全球气候变暖背景下,中国园艺植物生长季的农业气候资源发生变化,尤其是极端高温存在年际间波动性改变,直接影响了植物的生长和生物学性状的形成[8,23],高温胁迫对园艺植物生长的影响备受关注[15,24]。高温胁迫能激活细胞质内一系列的信号转导途径,改变膜脂结构和蛋白质结构,使细胞膜通透性发生改变,从而导致细胞内电解质外渗不断增加,电导率逐渐增大[14]。大量研究表明,电导率是反映细胞膜受损程度,评价植物耐热性的重要指标,许多遭受高温胁迫的植物,如绣球(Hydrangea macrophylla)、凌霄(Campsis grandiflova)、杜鹃(Rhododendron spp.)等植物叶片的电导率随着胁迫温度的增加而上升[15,20,21]。电导率法配合Logistic方程可计算出植物的LT50,能够简单、准确、快速地判定植物对高温的耐受性[15,20,21]。
本研究发现,随着胁迫温度的逐渐升高,牡丹叶片的电导率逐渐增大,对于ZB1而言,其耐热能力在不同温度梯度胁迫下均表现较弱,说明ZB1在高温胁迫条件下的自主调节能力较弱。此外,同一品种的耐热能力在不同温度梯度胁迫下存在差异,凤丹白在30~45 ℃细胞伤害率增加缓慢,耐热性较好,当温度继续上升>45 ℃时,凤丹白细胞伤害率上升较快,耐热性变弱。这一现象在植物中普遍存在,周媛等[25]发现在相对较低的高温胁迫下,金叶佛甲草的耐热能力较强,而在≥45 ℃的高温胁迫下,金叶佛甲草的耐热能力迅速降低,但胭脂红景天却与之相反。凌瑞等[15]发现,在35~55 ℃花手鞠的电导率细胞伤害率增长最慢,耐热性较强,而在55~65 ℃花手鞠的细胞质外渗最为严重,耐热性较弱。表明同一品种对不同强度高温胁迫的耐受程度及生理响应具有一定差异。本研究发现随着胁迫温度的不断升高,13个牡丹品种叶片的细胞伤害率呈一致性变化趋势,即缓慢上升-急剧上升-缓慢上升的 S形曲线增加趋势,通过显著性检验,符合Logistic方程。为进一步综合评价不同牡丹品种的耐热能力,计算13个牡丹品种的LT50,并通过有序样品最优分割聚类法,将13个牡丹品种按耐热能力强弱分为3类,B24、太平红、B10、凤丹荷、凤丹白的LT50均大于56.90 ℃,属于耐热品种(R),具有一定的高温适应能力,适合在高温地区推广应用;而泼墨紫、ZB1、海黄的LT50均小于54.81 ℃,属于不耐热品种(S),更适合种植于温度较低的温带地区。
本研究测定了叶片的相对电导率,所得到的LT50可有效评估牡丹的耐热能力。然而,牡丹的耐热性受多种因素的影响,其耐热生理过程错综复杂。有研究表明细胞膜相对通透性与植物耐热能力存在相关性,是评价植物耐热性的重要指标[15,20]。若要更加精确、全面地评价植物耐热性,需要结合叶绿素含量、叶绿素荧光参数、脯氨酸含量、超氧化物歧化酶活性、亚细胞结构等其他生理生化指标进行综合评价[14,26]。
参考文献:
[1] ZHOU S L,ZOU X H,ZHOU Z Q,et al. Multiple species of wild tree peonies gave rise to the ‘king of flowers, Paeonia suffruticosa Andrews[J].Proceedings of the royal society B,2014,281(1797): 20141687.
[2] 成仿云,李嘉珏.中国牡丹的输出及其在国外的发展Ⅰ:栽培牡丹[J].西北师范大学学报(自然科学版),1998(1):112-119.
[3] 李秀丽,陈法志,戢小梅,等.湖北省油用牡丹低产原因及增产关键栽培技术[J].湖北农业科学,2019,58(16):84-88,195.
[4] 张晓晓,牛立新,张延龙.中国芍药属牡丹组植物地理分布修订[A].中国园艺学会观赏园艺专业委员会,国家花卉工程技术研究中心.中国观赏园艺研究进展2017[C].2017.1-12.
[5] 徐 艳.几个牡丹品种的耐湿热生理生化特性研究[D].长沙:湖南农业大学,2007.
[6] 贺 丹,高小峰,吕博雅,等.牡丹、芍药花芽分化的形态学研究[J].河南农业科学,2014,43(12):117-120,124.
[7] FISCHER E M,SIPPEL S,KNUTTI R. Increasing probability of record-shattering climate extremes[J].Nature climate change,2021,11:689-695.
[8] 杨 阳,赵 娜,岳天祥.1980—2018年中国极端高温事件时空格局演变特征[J].地理科学,2022,42(3):536-547.
[9] 韩 欣,成仿云,肖佳佳,等.以‘凤丹白为母本的杂交及其育种潜力分析[J].北京林业大学学报,2014,36(4):121-125.
[10] 骞光耀,孔祥生,张淑玲.3个牡丹品种对高温胁迫的生理响应[J].江苏农业科学,2017,45(12):103-105.
[11] ZHANG Y Z,CHENG Y W,YA H Y,et al. Identification of heat shock proteins via transcriptome profiling of tree peony leaf exposed to high temperature[J].Genetics and molecular research,2015,14(3):8431-8442.
[12] 郝力慧,董 彬,朱绍华,等.牡丹响应高温胁迫的转录组分析及PsHSP基因表达[J].浙江农林大学学报, 2021,38(4):802-811.
[13] 朱绍华,马 进,郝力慧,等.江南牡丹叶片耐热差异蛋白的鉴定与分析[J].分子植物育种,2021,19(2):419-431.
[14] WAHID A,GELANI S,ASHRAF M,et al. Heat tolerance in plants: An overview[J].Environmental and experimental botany,2007,61:199-223.
[15] 凌 瑞,董 钠,刘惠斌,等.应用Logistic方程测定八个绣球品种的耐热性[J].北方园艺,2021(11):63-69.
[16] 李 进,顾 绘,殷琳毅.6个草莓品种高温半致死温度与耐热性评价[J].中国果树,2021(1):56-58.
[17] 李 进.矮壮素对小白菜高温半致死温度及耐热性的影响[J].中国瓜菜,2021(1):69-71.
[18] YU K D,ZHU K L,YE M J,et al. Heat tolerance of highbush blueberry is related to the antioxidative enzymes and oxidative protein-repairing enzymes[J].Scientia horticulturae,2016,198:36-43.
[19] ZHENG Y L,LI W Q,SUN W B. Effects of acclimation and pretreatment with abscisic acid or salicylic acid on tolerance of Trigonobalanus doichangensis to extreme temperatures[J].Biologia plantarum,2015,59(2):382-388.
[20] 袁雪涛,夏 冬,许永利,等.应用Logistic方程测定四种藤本植物的耐热性[J].北方园艺,2017(14):99-102.
[21] 刘婉迪,王 威,谢 倩,等.9个杜鹃品种的高温半致死温度与耐热性评价[J].西北林学院学报,2018,33(5):105-110.
[22] 朱根海,刘祖祺,朱培仁.利用Logistic方程确认植物组织低温半致死温度[J].南京农业大学学报,1986(3):11-16.
[23] 郭世博,张方亮,张镇涛,等.全球气候变暖对中国种植制度的可能影响XIV.东北大豆高产稳产区及农业气象灾害分析[J].中国农业科学,2022,55(9):1763-1780.
[24] WU Y Q,ZHAO D Q,HAN C X,et al. Biochemical and molecular responses of herbaceous peony to high temperature stress[J].Canadian journal of plant science,2016,96:474-484.
[25] 周 媛,郭彩霞,董艳芳,等.9种景天属轻型屋顶绿化植物的耐热性研究[J].西北农林科技大学学报(自然科学版),2014, 42(9):119-136.
[26] 陈志伟,李秀丽,戢小梅,等.武汉地区猕猴桃栽培品种耐热性评价[J].湖北农业科学,2022,61(13):69-72.
猜你喜欢
学生天地(2020年2期)2020-08-25 09:03:00
福建林业科技(2019年1期)2019-04-10 03:15:44
现代园艺(2017年13期)2018-01-19 02:28:13
江苏农业科学(2017年10期)2017-07-21 13:45:32
连环画报(2017年1期)2017-07-13 09:01:55
铝加工(2017年1期)2017-03-07 00:53:33
北方音乐(2016年12期)2016-08-23 03:20:03
中国塑料(2015年6期)2015-11-13 03:02:52
江苏农业科学(2015年7期)2015-08-20 11:59:23
质量与标准化(2015年11期)2015-07-21 14:54:29
