
巢湖西半湖真核浮游植物群落动态与理化因子相关性探究
2024-06-20 21:26:59赵冰冰朱鑫昊张华王金花黄健奚姗姗罗涛彭炜
南京信息工程大学学报 2024年3期
关键词:高通量测序
赵冰冰 朱鑫昊 张华 王金花 黄健 奚姗姗 罗涛 彭炜

摘要:浮游植物作为湖泊生态系统物质循环、能量循环的主要参与者,获取其群落结构信息对于调节湖泊生态系统具有重要意义.通过高通量测序检测巢湖西半湖冬季和夏季真核浮游植物的群落特征,在研究期间共检测出真核浮游植物7门71属,其中,夏季7门59属、冬季5门27属,以绿藻门、硅藻门为主,冬季较夏季优势属变化较大.夏季、冬季Shannon-Wiener指数均值分别为1.83、1.88,Pielou指数均值分别为0.75、0.83.两种多样性指数均值均为夏季小于冬季.水质分析结果表明,在研究期间巢湖西半湖水体TN、TP偏高,夏季、冬季水体理化指标具有显著性差异(P≤0.05).冗余分析表明,PO-4-P、TN、TP和NH+4-N浓度对真核浮游植物群落解释率较大,其中,PO-4-P浓度的影响达到显著水平(P≤0.05).Mantel相关性分析表明,真核浮游植物丰度与水温、溶解氧、pH、NH+4-N、TN和Chl.a有着良好的相关性.通过变差分解分析得到,季节因素对真核浮游植物群落解释度最高.
关键词高通量测序;巢湖西半湖;真核浮游植物;理化因子
中图分类号X171
文献标志码A
0引言
浮游植物是水生态系统中主要生产者之一,浮游植物的群落结构和动态变化可以深刻影响水生态系统和能量循环[1-2].浮游植物具有对环境变化反应迅速的特点,其丰度可以作为反映水体状况的重要生物指标[3].生物因素、气候因素、水文因素和地理因素等均会影响浮游植物群落结构的组成以及多样性[4-6].巢湖西半湖临近合肥市区,大量工业和城市废水流入西半湖,使巢湖富营养化程度从东向西逐渐增强,近一二十年间巢湖水华频率和严重程度有所增加[7].氮磷等营养元素与水温均会对浮游植物群落结构产生影响,且环境因子之间存在相互作用,这使得对浮游植物的调控变得更为复杂[8-9].分析湖区浮游植物群落组成的季节和空间分布及其与环境因子之间的关系,可为污染控制和水质评价提供科学依据.目前,对巢湖浮游植物群落与水质指标间的关系已有相关研究,但研究区域相对单一[10-11].本研究对巢湖河湖交汇处、岛屿、湖心、城镇和乡村周边水域均进行采样检测,在保证研究数据的准确与全面的基础上,探究巢湖西半湖水体理化因子与浮游植物群落之间的关系.
高通量测序法可以提供相关浮游植物群落组成的详细信息[12],较之通过显微镜进行的相关检测,更容易区分小型浮游植物并识别计数[13].本文采用高通量测序的方式检测浮游植物群落结构.首先对巢湖西半湖水体理化因子和浮游植物丰度的时空变化进行研究,并分析水体理化因子中对该区域真核浮游植物时空动态变化的影响.研究结果对日后在实验室中模拟水体理化因子中的某一种、某几种因素对真核浮游植物群落变化的影响有积极意义,也为巢湖水质保护积累了基础数据.期望能对研究区域门、属水平的真核浮游植物进行调控,为巢湖的环境保护及科学管理提供理论支撑.
1材料与方法
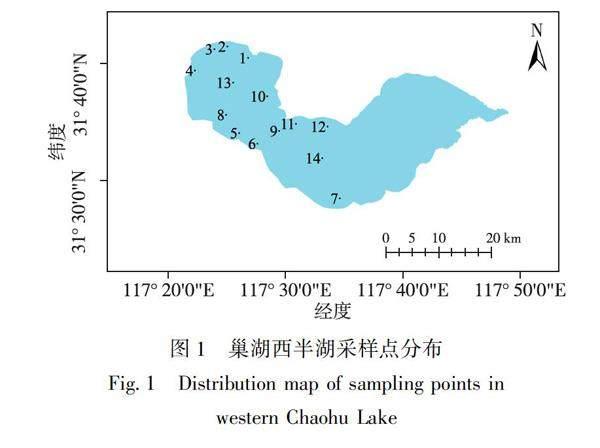
1.1采样点设置
本研究共设置14个采样点,具体布置如图1所示.采样点1~7分别对应南淝河、十五里河、塘西河、派河、杭埠河、白石天河、兆河与巢湖西半湖交汇区域;采样点8、9分别代表孤山和峔山岛湖区;采样点10代表受村落影响的湖区;采样点11、12代表受城镇影响的湖区;采样点13、14代表巢湖西半湖湖区水域中心.
1.2样品的采集
采样时间为2022年8月和2023年2月,使用玻璃采水器在采样点水面以下0.5m处采集水样2L,此水样用于检测巢湖水质指标.另使用玻璃采水器在水面以下0.5m处采集水样,水样经网孔直径为64μm的25号浮游生物滤网现场过滤,过滤浓缩后的水样装入离心管中,放置于保温箱内带回实验室.水样在实验室经0.22μm醋酸纤维滤膜过滤,收集水样中的真核浮游植物,通过高通量测序,鉴定真核浮游植物群落结构.滤膜放入离心管中,保存在液氮罐内.
1.3理化因子的检测
使用多参数水质分析仪在采样现场测得水温(WT)、酸碱度(pH)、溶解氧(DO).采集水样装入聚乙烯采样瓶中,避光保存,带回实验室.在实验室测得氨氮(NH+4-N)、硝氮(NO-3-N)、总氮(TN)、总磷(TP)、磷酸盐(PO-4-P)以及叶绿素a(Chl.a)浓度指标.样品的保存与测定均参照《水与废水监测分析方法》[14].
1.4浮游生物DNA提取与测序
使用TGuideS96磁珠法DNA提取试剂盒完成核酸的提取.使用酶标仪(synergyHTX)对提取的核酸进行浓度检测,后续进行PCR扩增,扩增后的PCR产物使用1.8%(质量分数)的琼脂糖进行电泳检测.PCR扩增及测序基于18SrDNA中V4区间,F:CCAGCASCYGCGGTAATTCC和R:ACTTTCGTTCTTGATYRA作为引物,扩增体系为10μL.扩增程序为:95℃预变性5min,25个循环(95℃变性30s,50℃退火30s,72℃延伸40s),最后72℃延伸7min.构建好的文库使用IlluminaNovaSeq6000(NovaSeq6000,Illumina)上机测序.
1.5测序控制与数据分析
首先使用Trimmomatic0.33软件,对测序得到的原始序列(RawReads)进行过滤,然后使用cutadapt1.9.1软件进行引物序列的识别与去除,再使用USEARCH10.0对双端序列(Reads)进行拼接并去除嵌合体,最终得到高质量序列(CleanReads)用于后续分析.使用USEARCH10.0在相似性97%的水平上对序列进行聚类,得到最佳分类单元(OperationalTaxonomicUnits,OTUs).将测序结果与Silva138数据库进行对比,使用朴素贝叶斯分类器对特征序列进行分类学注释,得到每个特征序列对应的物种分类信息.
获取足够的OTUs后,使用R4.3.1软件计算真核浮游植物群落的α多样性指数,包含Shannon-Wiener指数和Pielou指数,分析各点真核浮游植物多样性.此外,多样性指数可用于水体水质状况的判断,具体如表1所示.计算优势度指数(y′):
用主成分分析(PCA)来探究基于真核浮游植物群落结构和水体理化因子的样点分布特征,PCA分析和PERMANOVA分析基于R4.3.1软件vegan完成.使用Canoco5软件对真核浮游植物相对丰度进行消除趋势对应分析(DetrendedCorrespondenceAnalysis,DCA),以此结果来选择分析真核浮游植物纲分类水平下相对丰度和水体理化因子关系的方法:若第一轴长度<3,选择冗余分析(RedundancyAnalysis,RDA);若第一轴长度>4,选择典型相关分析(CanonicalCorrelationAnalysis,CCA);若3≤第一轴长度≤4,两者均可.Mantel相关分析探究水体理化因子对真核浮游植物门分类水平下的影响,基于R4.3.1软件vegan、ggcor、dplyr、ggplot2包完成.变差分解分析(VariancePartitioningAnalysis,VPA)基于R4.3.1软件vegan包完成.采样点图使用ArcMap10.7完成绘制.
2结果与分析
2.1巢湖水体理化因子分析
巢湖西半湖水体理化因子变化如图2所示,水体理化因子具有时空差异性.水温(WT)随季节变化差异较大,最大差值27℃.夏季溶解氧(DO)整体低于冬季,范围在6.80~9.73mg·L-1.巢湖水体整体呈弱碱性,最低pH为8.18,夏季pH整体略高于冬季,夏季、冬季均值分别为8.57、8.38.冬季NH+4-N质量浓度均值0.13mg·L-1,低于夏季均值0.65mg·L-1.除采样点13外,TN质量浓度夏季大于冬季,而且夏季有7个采样点TN指标超过地表水Ⅴ类的标准.夏季、冬季NO-3-N质量浓度范围分别为0.64~1.34和0.35~1.56mg·L-1.TP质量浓度偏高,夏季、冬季分别有8个和9个采样点达到地表水Ⅴ类的标准,夏季、冬季总体差异较小.PO-4-P质量浓度随季节波动较小,但夏季采样点2、3和其他采样点有一定差异.Chl.a质量浓度夏季波动较大,范围在11.41~238.46μg·L-1,夏季、冬季均值分别为61.13和12.48μg·L-1.
采用PCA分析各采样点水体理化因子的相似度,两点坐标直线距离越近,则两点的水体理化因子差异越小.PCA分析结果(图3)显示,PCA两轴共解释了69.3%,所有采样点水质理化因子均在95%置信区间内,经过PERMANOVA分析,巢湖西半湖夏季、冬季水体理化因子具有显著性差异(P≤0.05).夏季采样点均在一四象限,冬季采样点均在二三象限,所有采样点在PCA轴1具有很好的区分度,TN、TP、WT等水体理化因子与PCA轴1具有正相关关系,仅DO与PCA轴1具有负相关关系.
2.2巢湖西半湖真核浮游植物群落组成
2.2.1测序结果
对夏季、冬季共28个样品18SrDNA的V4区间扩增测序后,共获得2235345对Reads,双端Reads质控、拼接后共产生1953378条CleanReads,每个样品至少产生51468条CleanReads.本研究共检测出7门71属真核浮游植物.夏季检测出7门59属,其中,绿藻门24属、硅藻门15属、金藻门10属、隐藻门4属、甲藻门4属、黄藻门定鞭藻门各1属;冬季检测出5门27属,其中,绿藻门15属、硅藻门4属、金藻门4属、甲藻门3属、黄藻门1属.冬季除黄藻门之外,其他各藻门包含藻属均有不同程度减少,减少最多的为硅藻门,共减少11属,夏季、冬季真核浮游植物群落结构在属水平上差异较大,夏季、冬季均检出的真核浮游植物仅15属.
2.2.2真核浮游植物门水平组成及变化
夏季、冬季两次采样真核浮游植物门分类水平下相对丰度如图4所示,研究区域真核浮游植物相对丰度占比较高的均为绿藻门和硅藻门.夏季、冬季绿藻门相对丰度均值分别为55.78%和54.85%,硅藻门则分别为24.92%和27.71%.与冬季相比,夏季各采样点绿藻门、硅藻门相对丰度差异较大.夏季绿藻门相对丰度在24%~86%,相对丰度较低的1、3、4号采样点,均在巢湖西北部,夏季硅藻门相对丰度在6%~70%;冬季各采样点绿藻门、硅藻门相对丰度分别在31%~78%和16%~38%,与夏季相比波动较小.夏季甲藻门相对丰度最大采样点仅为5%,冬季甲藻门相对丰度明显增加,各采样点甲藻门平均丰度为13.85%.同时,隐藻门、定鞭藻门在冬季均未检出.
2.2.3优势属的变化
研究期内优势属如表2所示,巢湖西半湖优势属主要由绿藻门、硅藻门和隐藻门构成,夏季和冬季优势属有较大差异.衣藻属(Chlamydomonas)在夏季优势度最大,但在冬季却未检出,沟链藻属(Aulacoseira)、角毛藻属(Chaetoceros)、隐藻属(Cryptomonas)也有类似现象.齿状藻属(Odontella)在夏季未检出,但在冬季成为优势属之一;麦可属(Mychonastes)在夏季虽有检出,但优势度很低,在冬季却成为优势度最大的藻属.仅绿藻纲未定属(unclassified_Chlorophyceae)为夏季、冬季共同优势属.
2.2.4真核浮游植物时空相似性比较
通过PCA对巢湖西半湖夏季、冬季真核浮游植物群落结构进行比较分析,结果如图5所示.结果表明,PCA轴1的解释率为24.4%,PCA轴2解释率为16.2%,总解释率为40.6%.冬季各采样点真核浮游植物群落分布在二三象限,夏季各采样点真核浮游植物群落则主要分布在三四象限.通过PERMANOVA分析可知,巢湖西半湖夏季、冬季真核浮游植物群落结构具有显著差异性(R2=0.338,P=0.001).
2.2.5多样性分析
夏冬季巢湖西半湖真核浮游植物多样性指数有一定差异,结果如表3所示.夏季Shannon-Wiener指数的范围在1.04~2.52,均值为1.83,冬季Shannon-Wiener指数的范围在1.56~2.28,均值为1.88,如按Shannon-Wiener指数均值评价水质,巢湖西半湖夏季、冬季整体处于中污染状态.夏季Pielou指数范围在0.53~0.91,均值为0.75,冬季Pielou指数范围在0.76~0.89,均值为0.83,如按Pielou指数均值评价水质,巢湖西半湖夏季整体偏轻污染状态,冬季整体偏清洁状态.巢湖西半湖夏季Shannon-Wiener指数、Pielou指数均值均小于冬季,故推测夏季水生态系统稳定性整体劣于冬季.结果显示,Pielou指数对于研究区内水质情况有一定区分能力,但Shannon-Wiener指数对于研究区内水质情况区分能力较差.
2.3水体理化因子与真核浮游植物的关系
2.3.1水体理化因子与真核浮游植物纲水平RDA分析
选取NH+4-N、TN、TP、PO-4-P、Chl.a质量浓度等9个水体理化因子指标,与绿藻纲、共球藻纲、隐藻纲等真核浮游植物纲水平相对丰度进行DCA分析.第1轴长度为1.21,故选择进行RDA分析,结果如图6所示.第1轴和第2轴解释量分别为44.3%和4.58%.环境因子中PO-4-P、TN、TP和NH+4-N质量浓度对真核浮游植物群落解释率较大,其中,PO-4-P质量浓度的影响达到显著水平(P≤0.05).共球藻纲、小豆藻纲、硅藻纲、隐藻纲、金藻纲和间藻纲均在第一象限,与NH+4-N、TN、TP、Chl.a质量浓度和水温均呈正相关.溶解氧、PO-4-P质量浓度和各纲均呈负相关.
2.3.2水体理化因子与真核浮游植物门分类水平下相对丰度Mantel相关性分析
为进一步了解水体理化因子对真核浮游植物的影响,将真核浮游植物门分类水平下的相对丰度与水体理化因子进行Mantel相关性分析,结果如图7所示.各藻门与WT、DO、pH以及NH+4-N、TN和Chl.a质量浓度有着良好的相关性.其中,绿藻门与水温,溶解氧,NH+4-N、TN质量浓度呈极显著正相关(P<0.01);隐藻门、硅藻门与水温,溶解氧,pH,NH+4-N、TN、Chl.a质量浓度呈极显著正相关(P<0.01);金藻门与NH+4-N、TN、Chl.a质量浓度呈极显著正相关(P<0.01);甲藻门与水温、溶解氧、NH+4-N质量浓度呈极显著正相关(P<0.01).
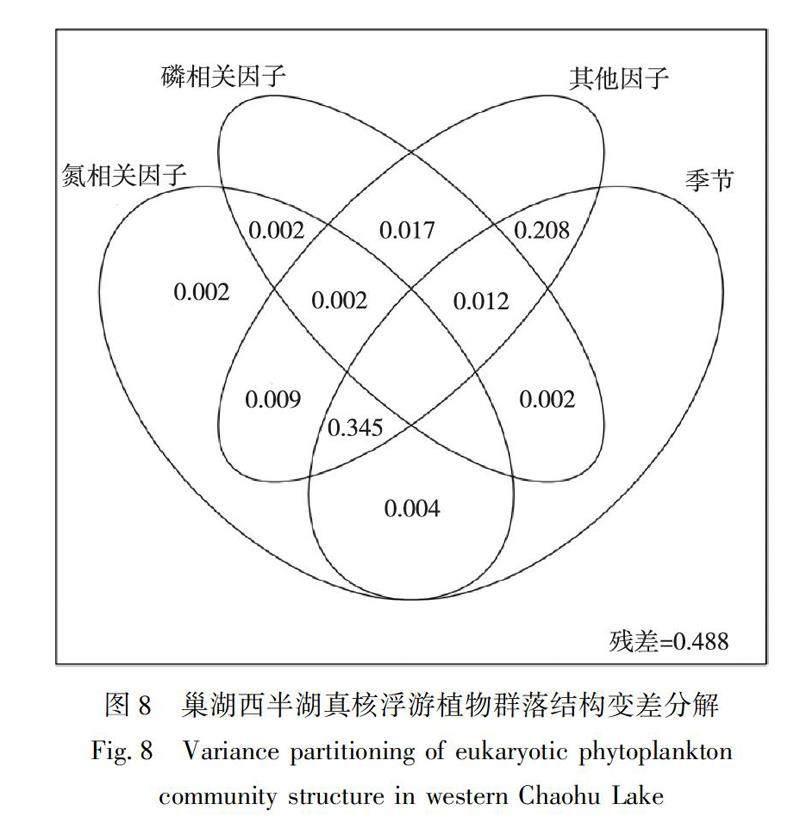
2.3.3真核浮游植物影响因子变差分解
为了解不同类型的水体理化因子和季节对巢湖西半湖真核浮游植物群落结构的影响,对氮相关因子(包含NH+4-N、TN、NO-3-N)、磷相关因子(TP、PO-4-P)、其他因子(水温、溶解氧、pH、Chl.a)和季节进行变差分解,结果如图8所示.这4类因子共解释了60.3%的真核浮游植物群落结构变化.其中,季节因素解释率最高,氮相关因子与季节共同解释率为20.8%,氮相关因子、其他因子与季节共同解释率为34.5%.
3讨论
3.1巢湖西半湖真核浮游植物丰度变化
本次研究通过高通量测序技术共检测出真核浮游植物7门71属,其中夏季检出7门59属、冬季检出5门27属,较显微镜检测结果更为丰富[17].检出真核浮游植物主要为绿藻门、硅藻门、金藻门、隐藻门和甲藻门,其中,绿藻门藻属数量最多(夏季占40.7%,冬季55.6%),这与前人对巢湖的调查结果基本一致,在裸藻门结果中略有偏差,结果可能是和本次选取18SrDNA中V4区间引物的特异性有关[18].本研究结果中,隐藻门生物在夏季占有一定比例,但在冬季未检出,这种情况可能因为隐藻门与水温有正相关的关系,冬季水温下降,导致隐藻门生物在冬季没有被检出[19].目前一般认为硅藻门生物喜低温,而绿藻门生物更适合温度较高的水体,有学者对三峡水库上游长寿湖和南海湖进行研究,冬季硅藻门相对丰度均高于绿藻门相对丰度[20-21].但本研究结果却显示,冬季巢湖西半湖绿藻门相对丰度大于硅藻门相对丰度,导致这一结果的原因可能是对真核浮游植物丰度的检测方法不同,高通量测序可以弥补显微镜观测可能丢失的信息,如所检出冬季第一优势属绿藻门麦可属(Mychonastes)由于粒径较小,难以通过显微镜观测[22].张春霞等[23]通过高通量测序的方式对丹江口水库真核浮游植物丰度进行检测,也出现冬季绿藻门相对丰度大于硅藻门相对丰度的现象.浮游植物属水平下,只有一属在夏季、冬季均达到优势属的水平,其他单一季节优势属只在相应季节达到优势属的水平,在其他季节不满足优势属条件,甚至未检出.Shi等[24]发现,在特定的时间空间下只有少数物种主导着浮游植物群落结构,同时伴随着很多相对丰度较低的物种,但这些相对丰度较低的物种,一年仍有至少1~2次机会达到高丰度,这与优势属变化较大相契合.
3.2巢湖水质状况与水质评价
根据检测结果发现,巢湖水质具有时空差异性,巢湖西半湖靠近合肥市区,使其接收到更多的污染物,致使巢湖西半湖富营养化程度大于东半湖.结果表明,靠近合肥市区的采样点(1~4号)TN、NH+4-N质量浓度均值偏高.不同季节巢湖水质有一定差异,虽然冬季和夏季TP、PO-4-P相近,但冬季溶解氧含量明显高于夏季,TN、NH+4-N质量浓度均值却低于夏季,故认为巢湖冬季水质优于夏季.本实验夏季、冬季两次采样,TN质量浓度均值1.69mg·L-1,TP质量浓度均值0.10mg·L-1,相对前期研究结果较低,推测巢湖治理效果逐渐显现[25].
群落多样性指数是反映群落结构稳定性的重要参数,本研究选用了Shannon-Wiener指数和Pielou指数进行对浮游植物群落表征.其中,Shannon-Wiener指数夏季、冬季范围分别为1.04~2.52、1.56~2.28,Pielou指数夏季、冬季范围分别为0.53~0.91、0.76~0.89.两种多样性指数均是夏季波动大于冬季,间接表明夏季不同水域真核浮游植物群落结构差异大于冬季,这与夏季绿藻门、硅藻门波动大于冬季相契合.一般情况下,浮游植物的多样性指数越大,认为其群落结构多样性越强,稳定性越高,水质条件越好[26].但近年来也有学者在研究中发现浮游植物多样性指数在评价水质中存在不准确的现象,本次研究分析了两种浮游植物多样性指数对研究区域水质评价的适用性.对于巢湖西半湖不同季节水体理化因子进行分析,得到夏季、冬季水体理化因子具有显著性差异,但结果中Pielou指数对研究区域内夏季冬季水体有区分能力,而Shannon-Wiener指数对研究区域内水体区分能力较差.有学者认为,在大面积湖泊中经度纬度的梯度变化以及显著的区域特征会给浮游植物多样性指数带来很大的干扰[27].也有学者指出,浮游植物多样性指数除与水质相关外,气候也会对其产生深远影响[28].在诸如河流、湖泊等地理跨度大、地形复杂的生态系统中,多样性指数能在一定程度上表现生态环境变化,但对水质评价尚需要结合检测所得水质指标进行综合考虑[29].
3.3巢湖西半湖浮游植物与影响因素
浮游植物群落结构与水体理化因子的关系一直是水生态研究的重点之一,但是不同时间或相同时间的不同区域浮游生物群落结构的影响因子各不相同.本研究通过RDA分析和Mantel分析,显示浮游植物相对丰度和水体理化因子有着很好的相关性.水温一般被认为是影响浮游植物群落结构的关键因素,水温通过调节同化作用、细胞内酶的活性以及代谢速率的方式,直接或间接影响浮游植物的群落结构[30-31].真核浮游植物生长速率受到水温影响,许多浮游植物在水温20℃左右,可以达到最大生长速率[32].pH作为一种重要的水体理化指标,不同真核浮游植物生长繁殖对水体pH有着不同的要求[33].通过RDA分析,pH与绿藻门共球藻纲呈正相关,但与绿藻门绿藻纲呈负相关.调查期间,巢湖西半湖水体整体偏碱性,碱性水体有利于浮游植物吸收CO2,从而进行光合作用[34].N、P元素是真核浮游植物生长代谢必需的元素.有学者认为氮磷比(TN/TP)是营养盐结构的一个重要指标,氮磷比>10为磷限制性状态,氮磷比<7为氮限制性状态[35].N元素不足,将会导致主要光合色素Chl.a的含量大幅下降,并增加非光合色素的含量;P元素不足,浮游植物增加无磷化合物的合成,导致其他化合物代替细胞膜中的磷脂[36].NH+4-N是能够被浮游植物直接吸收利用的氮形态,NH+4-N浓度增加,对浮游植物有促进作用[37].通过RDA分析得到,浮游植物各纲均与PO-4-P浓度呈负相关,出现磷元素对真核浮游有抑制的现象.前人通过NEB实验[38],发现添加磷可能出现抑制浮游植物生长的状况,与本次研究相契合.Chl.a浓度通常可以被用来表征浮游植物生物量[39].结果显示,夏季、冬季Chl.a质量浓度分别为61.13和12.48μg·L-1,夏季浮游植物生物量大于冬季,说明夏季更适合浮游植物生长.本研究结果将有利于之后模型的建立、运算以及推演,促进建立水体理化因子与真核浮游植物间的互相约束关系模型,为进一步研究或调控区域水质、浮游植物群落起到正向作用.
4结论
1)通过高通量测序,检测出巢湖西半湖夏季、冬季真核浮游植物共7门71属,其中,夏季7门59属,冬季5门27属.在门分类水平下,夏季、冬季相对丰度较高的均为绿藻门和硅藻门.夏季、冬季真核浮游植物群落结构具有显著差异性(P=0.001).
2)在研究期间,巢湖西半湖水体水质氮磷超标,夏季水质劣于冬季,夏季、冬季水质有显著差异性(P≤0.05).Shannon-Wiener指数和Pielou指数夏季波动大于冬季,均值夏季小于冬季,显示冬季群落结构相较于夏季更稳定.同时,Pielou指数对研究区域内水质有一定区分能力,Shannon-Wiener指数对研究区域内水质区分能力较差,对于巢湖西半湖水质,建议采样生物评价结合理化指标等多种方法综合评价.
3)RDA分析结果表明,PO-4-P、TN、TP和NH+4-N对真核浮游植物群落解释率较大.其中,PO-4-P的影响达到显著水平(P≤0.05).Mantel相关性分析结果表明,真核浮游植物与WT、DO、pH、NH+4-N、TN和Chl.a有着良好的相关性.变差分解分析结果表明,季节因素对真核浮游植物群落解释度最高.
参考文献
References
[1]
ZhangHH,YangYS,LiuX,etal.Novelinsightsinseasonaldynamicsandco-existencepatternsofphytoplanktonandmicro-eukaryotesindrinkingwaterreservoir,NorthwestChina:DNAdataandecologicalmodel[J].ScienceoftheTotalEnvironment,2023,857:159160
[2]McNabneyDWG,MangalV,KirkwoodAE,etal.PhytoplanktonmetaboliteprofilesfromtwoLakeOntarioareasofconcernrevealdifferencesassociatedwithtaxonomiccommunitycomposition[J].ScienceoftheTotalEnvironment,2023,871:162042
[3]TangCH,YiYJ,YangZF,etal.Planktonicindicatorsoftrophicstatesforashallowlake(BaiyangdianLake,China)[J].Limnologica,2019,78:125712
[4]SeverianoJDS,Almeida-MeloVLDS,Bittencourt-OliveiraMDC,etal.Effectsofincreasedzooplanktonbiomassonphytoplanktonandcyanotoxins:atropicalmesocosmstudy[J].HarmfulAlgae,2018,71:10-18
[5]ChangC,GaoL,WeiJ,etal.Spatialandenvironmentalfactorscontributingtophytoplanktonbiogeographyandbiodiversityinmountainpondsacrossalargegeographicarea[J].AquaticEcology,2021,55(2):721-735
[6]JiangYJ,HeW,LiuWX,etal.TheseasonalandspatialvariationsofphytoplanktoncommunityandtheircorrelationwithenvironmentalfactorsinalargeeutrophicChineselake(LakeChaohu)[J].EcologicalIndicators,2014,40:58-67
[7]FanYY,LiBB,YangZC,etal.MediationoffunctionalgeneandbacterialcommunityprofilesinthesedimentsofeutrophicChaohuLakebytotalnitrogenandseason[J].EnvironmentalPollution,2019,250:233-240
[8]LiuXM,ChenLW,ZhangGX,etal.Spatiotemporaldynamicsofsuccessionandgrowthlimitationofphytoplanktonfornutrientsandlightinalargeshallowlake[J].WaterResearch,2021,194:116910
[9]RaoK,ZhangX,WangM,etal.Therelativeimportanceofenvironmentalfactorsinpredictingphytoplanktonshiftingandcyanobacteriaabundanceinregulatedshallowlakes[J].EnvironmentalPollution,2021,286:117555
[10]王徐林,张民,殷进.巢湖浮游藻类功能群的组成特性及其影响因素[J].湖泊科学,2018,30(2):431-440
WANGXulin,ZHANGMin,YINJin.CompositionandinfluentialfactorsofphytoplanktonfunctiongroupsinLakeChaohu[J].JournalofLakeSciences,2018,30(2):431-440
[11]路娜,尹洪斌,邓建才,等.巢湖流域春季浮游植物群落结构特征及其与环境因子的关系[J].湖泊科学,2010,22(6):950-956
LUNa,YINHongbin,DENGJiancai,etal.SpringcommunitystructureofphytoplanktonfromLakeChaohuanditsrelationshiptoenvironmentalfactors[J].JournalofLakeSciences,2010,22(6):950-956
[12]ChenZF,ZhangQC,KongFZ,etal.Resolvingphytoplanktontaxabasedonhigh-throughputsequencingduringbrowntidesintheBohaiSea,China[J].HarmfulAlgae,2019,84:127-138
[13]SimonM,López-GarcíaP,DeschampsP,etal.Markedseasonalityandhighspatialvariabilityofprotistcommunitiesinshallowfreshwatersystems[J].TheISMEJournal,2015,9(9):1941-1953
[14]国家环境保护总局.水和废水监测分析方法[M].4版.北京:中国环境科学出版社,2002
[15]杜红春,朱挺兵,龚进玲,等.澜沧江西藏段浮游植物群落结构特征及其与环境因子的关系[J].生态学杂志,2022,41(9):1769-1777
DUHongchun,ZHUTingbing,GONGJinling,etal.PhytoplanktoncommunitystructureanditsrelationshipwithenvironmentalfactorsintheXizangreachesoftheLancangRiver[J].ChineseJournalofEcology,2022,41(9):1769-1777
[16]CavalcantiLF,FeitosaFADN,CutrimMVJ,etal.DriversofphytoplanktonbiomassanddiversityinamacrotidalbayoftheAmazonMangroveCoast,aRamsarsite[J].Ecohydrology&Hydrobiology,2022,22(3):435-453
[17]吴转璋,朱超,唐萍,等.巢湖湖区浮游植物群落与水质因子相关性分析[J].生物学杂志,2023,40(1):79-84
WUZhuanzhang,ZHUChao,TANGPing,etal.CorrelationanalysisofphytoplanktoncommunityandwaterqualityfactorsinChaohuLake[J].JournalofBiology,2023,40(1):79-84
[18]朱超,杨晓冉,赵彬,等.2017年夏季巢湖水华期间浮游植物与蓝藻毒素的时空变化特征[J].中国环境监测,2018,34(6):103-112
ZHUChao,YANGXiaoran,ZHAOBin,etal.SpatiotemporaldynamicsofphytoplanktonandcyanotoxinsinChaohuLakeduringsummercyanobacterialbloomsof2017[J].EnvironmentalMonitoringinChina,2018,34(6):103-112
[19]张俊芳,陈威,宋以兴,等.雅砻江下游浮游植物群落结构时空变化特征及环境驱动因子[J].长江流域资源与环境,2023,32(5):950-960
ZHANGJunfang,CHENWei,SONGYixing,etal.Spatio-temporalvariationofphytoplanktoncommunitystructureandenvironmentaldrivingfactorsinlowerreachesofYalongRiver[J].ResourcesandEnvironmentintheYangtzeBasin,2023,32(5):950-960
[20]雷波,刘朔孺,张方辉,等.三峡水库上游长寿湖浮游藻类的季节变化特征及关键环境影响因子[J].湖泊科学,2017,29(2):369-377
LEIBo,LIUShuoru,ZHANGFanghui,etal.SeasonalphytoplanktondynamicsanditsrelationshipwithkeyenvironmentalfactorsinLakeChangshou,upperreachesoftheThreeGorgesReservoir[J].JournalofLakeSciences,2017,29(2):369-377
[21]高静湉,毋凡琛,何桂豪,等.寒区城市湖泊浮游植物群落特征及环境驱动因子[J].环境科学与技术,2021,44(6):1-10
GAOJingtian,WUFanchen,HEGuihao,etal.Phytoplanktoncommunitycharacteristicsandenvironmentaldrivingfactorsofurbanlakesincoldregions[J].EnvironmentalScience&Technology,2021,44(6):1-10
[22]李艳晖,胡明明,沈银武,等.中国淡水绿藻纲新记录属:麦可属(Mychonastes)[J].水生生物学报,2013,37(3):473-480
LIYanhui,HUMingming,SHENYinwu,etal.Mychonastes,anewrecordedgenusoffreshwaterchlorophyceaeisolatedfromaChineselake[J].ActaHydrobiologicaSinica,2013,37(3):473-480
[23]张春霞,贺玉晓,郭晓明,等.丹江口水库夏冬季真核浮游植物群落结构变化及其驱动因素[J].河南理工大学学报(自然科学版),2022,41(6):110-122
ZHANGChunxia,HEYuxiao,GUOXiaoming,etal.CommunitystructurevariationsanddrivingfactorsofeukaryotesphytoplanktoninDanjiangkoureservoirinsummerandwinter[J].JournalofHenanPolytechnicUniversity(NaturalScience),2022,41(6):110-122
[24]ShiXL,LiSN,ZhangM,etal.TemperaturemainlydeterminesthetemporalsuccessionofthephotosyntheticpicoeukaryotecommunityinLakeChaohu,ahighlyeutrophicshallowlake[J].ScienceoftheTotalEnvironment,2020,702:134803
[25]袁强,余丽,李卫华,等.巢湖和太湖春季微囊藻毒素动态差异及与环境因子的关系[J].环境工程技术学报,2021,11(2):267-277
YUANQiang,YULi,LIWeihua,etal.DynamicdifferencesofmicrocystinsandtheirrelationshipwithenvironmentalfactorsinspringinLakeChaohuandLakeTaihu[J].JournalofEnvironmentalEngineeringTechnology,2021,11(2):267-277
[26]贺玉晓,郑永坤,李卫国,等.丹江口水库早春真核浮游植物群落结构特征及其与环境因子的关系[J].环境科学学报,2021,41(6):2192-2200
HEYuxiao,ZHENGYongkun,LIWeiguo,etal.CharacteristicsofeukaryoticphytoplanktoncommunitystructureinearlyspringanditsrelationshipwithenvironmentalfactorsinDanjiangkouReservoir[J].ActaScientiaeCircumstantiae,2021,41(6):2192-2200
[27]王世欢,张生,武蓉,等.寒旱区湖泊浮游植物特征及其对营养状态的指示作用[J].中国环境科学,2023,43(1):311-320
WANGShihuan,ZHANGSheng,WURong,etal.Characteristicsofphytoplanktonincoldandaridareasandtheirindicatoroftrophicstatus[J].ChinaEnvironmentalScience,2023,43(1):311-320
[28]BloisJL,WilliamsJW,FitzpatrickMC,etal.Spacecansubstitutefortimeinpredictingclimate-changeeffectsonbiodiversity[J].ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica,2013,110(23):9374-9379
[29]DingYT,PanBZ,ZhaoGN,etal.Geo-climaticfactorsweakentheeffectivenessofphytoplanktondiversityasawaterqualityindicatorinalargesediment-ladenriver[J].ScienceoftheTotalEnvironment,2021,792:148346
[30]钱玺亦,李金彪,敖文,等.呼伦湖浮游植物群落季节动态及其与环境因子的关系[J].湖泊科学,2022,34(6):1814-1827
QIANXiyi,LIJinbiao,AOWen,etal.SeasonaldynamicsofphytoplanktonanditsrelationshipwithenvironmentalfactorsinLakeHulun[J].JournalofLakeSciences,2022,34(6):1814-1827
[31]SuXM,SteinmanAD,XueQJ,etal.Temporalpatternsofphyto-andbacterioplanktonandtheirrelationshipswithenvironmentalfactorsinLakeTaihu,China[J].Chemosphere,2017,184:299-308
[32]AbirhireO,DaviesJM,ImtiazyN,etal.ResponseofphytoplanktoncommunitycompositiontophysicochemicalandmeteorologicalfactorsunderdifferenthydrologicalconditionsinLakeDiefenbaker[J].ScienceoftheTotalEnvironment,2023,856:159210
[33]KimH,SpivackAJ,Menden-DeuerS.pHalterstheswimmingbehaviorsoftheraphidophyteHeterosigmaakashiwo:implicationsforbloomformationinanacidifiedocean[J].HarmfulAlgae,2013,26:1-11
[34]胡愈炘,曹亮,屈仁超,等.长江流域着生藻类群落结构的空间格局及其生态评价[J].环境科学,2022,43(8):3998-4007
HUYuxin,CAOLiang,QURenchao,etal.CommunitycompositionandassessmentoftheaquaticecosystemofperiphyticalgaeintheYangtzeRiverBasin[J].EnvironmentalScience,2022,43(8):3998-4007
[35]宋勇军,戚菁,刘立恒,等.程海湖夏冬季浮游植物群落结构与富营养化状况研究[J].环境科学学报,2019,39(12):4106-4113
SONGYongjun,QIJing,LIULiheng,etal.EutrophicationstatusandphytoplanktoncommunitystructureinChenghaiLakeinsummerandwinter[J].ActaScientiaeCircumstantiae,2019,39(12):4106-4113
[36]JiangM,NakanoS.Thecrucialinfluenceoftrophicstatusontherelativerequirementofnitrogentophosphorusforphytoplanktongrowth[J].WaterResearch,2022,222:118868
[37]张琦,陈宇琛,林育青,等.澜沧江梯级水库浮游植物群落结构特征及其关键驱动因子[J].湖泊科学,2023,35(2):530-539
ZHANGQi,CHENYuchen,LINYuqing,etal.CharacteristicofphytoplanktoncommunitystructureanditsdrivingfactorsalongthecascadereservoirsintheLancangRiver[J].JournalofLakeSciences,2023,35(2):530-539
[38]李利强,黄代中,熊剑,等.洞庭湖浮游植物增长的限制性营养元素研究[J].生态环境学报,2014,23(2):283-288
LILiqiang,HUANGDaizhong,XIONGJian,etal.NutrientlimitingphytoplanktongrowthinDongtingLake[J].EcologyandEnvironmentalSciences,2014,23(2):283-288
[39]ZouW,ZhuG,XuH,etal.ElucidatingphytoplanktonlimitingfactorsinlakesandreservoirsoftheChineseeasternplainsecoregion[J].JournalofEnvironmentalManagement,2022,318:115542
Correlationbetweeneukaryoticphytoplanktoncommunitydynamics
andphysicochemicalfactorsinwesternChaohuLake,China
ZHAOBingbing1,2ZHUXinhao2ZHANGHua1,2WANGJinhua1,2
HUANGJian1,2XIShanshan1,2LUOTao2PENGWei2
1AnhuiInstituteofEcologicalCivilization,AnhuiJianzhuUniversity,Hefei230601,China
2SchoolofEnvironmentandEnergyEngineering,AnhuiJianzhuUniversity,Hefei230601,China
AbstractPhytoplanktonisamajorparticipantinthematerialandenergycyclesoflakeecosystems,andtheinformationofitscommunitystructureisofgreatsignificanceincopingwithandregulatinglakeecosystems.Inthisstudy,thecommunitycharacteristicsofeukaryoticphytoplanktoninthewesternChaohuLakeinwinterandsummerwereobtainedviahigh-throughputsequencing.Atotalof7phylaand71generaofeukaryoticphytoplanktonweredetectedduringthesurvey,including7phylaand59generainsummerand5phylaand27generainwinter,dominatedbyChlorophytaandBacillariophyta,andthedominantgeneravariedgreatlyinwintercomparedwiththoseinsummer.ThemeanvaluesofShannon-Wienerindexinsummerandwinterwere1.83and1.88respectively,andthePielouindexwereaveraged0.75and0.83forsummerandwinterrespectively.TheresultsofwaterqualityanalysisindicatedthatTNandTPwererelativelyhighinthewesternChaohuLakeduringthestudyperiod,andthephysicochemicalfactorsofthewaterbodyvariedsignificantlybetweensummerandwinter(P≤0.05).RedundancyanalysisshowedthattheeukaryoticphytoplanktoncommunitycanberoughlyexplainedbyPO-4-P,TN,TPandNH+4-N,especiallythePO-4-P(P≤0.05).MantelcorrelationanalysisshowedaclosecorrelationbetweeneukaryoticphytoplanktonabundanceandWT,DO,pH,NH+4-N,TNandChl.a.Variancepartitioninganalysisshowedthatseasonalfactorsexplainedmostoftheeukaryoticphytoplanktoncommunitydynamics.
Keywordshigh-throughputsequencing;westernChaohuLake;eukaryoticphytoplankton;physicochemicalfactors
猜你喜欢
江苏农业科学(2017年20期)2017-11-30 14:53:29
江苏农业科学(2017年16期)2017-10-27 14:31:12
湖北农业科学(2017年13期)2017-08-08 05:09:44
中国中药杂志(2017年13期)2017-07-31 08:28:44
山东工业技术(2017年12期)2017-07-06 18:29:27
中国中药杂志(2017年7期)2017-05-26 00:05:40
中国医药导报(2017年9期)2017-05-11 18:44:13
中国中药杂志(2016年24期)2017-04-18 17:42:52
中国中药杂志(2017年4期)2017-03-28 17:06:08
中国中药杂志(2017年3期)2017-03-20 21:11:11
