
基于Biomod2组合模型的我国山杨潜在分布区研究
2024-06-15 00:00:00高明龙铁牛张晨李凤滋乌雅瀚罗奇辉王子瑞刘磊萨如拉
南京林业大学学报(自然科学版) 2024年2期
收稿日期Received:2022-05-13""" 修回日期Accepted:2022-08-05
基金项目:内蒙古自治区科技计划项目(2020GG0067)。
第一作者:高明龙(gml9652@foxmail.com)。
*通信作者:萨如拉(sarula213@163.com),教授。
引文格式:
高明龙,铁牛,张晨,等. 基于Biomod2组合模型的我国山杨潜在分布区研究. 南京林业大学学报(自然科学版),2024,48(2):247-255.
GAO M L, TIE N, ZHANG C, et al. Modelling the potential distribution area of" Populus davidiana" in China based on the Biomod2. Journal of Nanjing Forestry University (Natural Sciences Edition),2024,48(2):247-255.
DOI:10.12302/j.issn.1000-2006.202205022.
摘要:【目的】通过探究环境变化对山杨(Populus davidiana)分布的影响,为山杨资源的保护和开发提供理论支撑。【方法】根据山杨的134条地理分布数据,结合18个气候、土壤及地形因子,基于Biomod2软件包构建组合模型,模拟我国山杨潜在分布区在未来3种气候条件模式下的空间分布格局变化,并确定影响山杨分布的主要环境变量。【结果】我国山杨当前潜在适生分布区主要位于400 mm等降水线两侧较高纬度或较高海拔地区,总面积约为1 560 340.9 km2,约占我国陆地面积的16.2%,其中大兴安岭、长白山、太行山、秦岭、祁连山南麓、横断山、云贵高原等地区为山杨高度适生区;在未来气候条件下,山杨适生区整体呈向西南方向收缩趋势,生境适宜度总体呈下降趋势;影响山杨分布主要环境变量为最热月最高气温、年降水量和海拔;基于5个最优单一模型构建的组合模型比单一模型对山杨适生区预测结果更好,训练集平均受试者工作特征曲线下面积和真实技巧统计值分布为 0.91和0.73,预测准确度较高。【结论】我国山杨空间分布格局主要受水热条件影响,海拔也是影响山杨分布的重要因素。在未来气候条件下,山杨分布区面积将随气候变暖的程度逐渐减少。以山杨作为用材林和生态公益林树种进行造林时,造林地点应选择未来生境适宜度变化不大的地区,以降低未来由于气候变化造成的损失。
关键词:山杨;Biomod2软件包;组合模型;潜在分布区;气候变暖
中图分类号:S792.114""""" 文献标志码:A开放科学(资源服务)标识码(OSID):
文章编号:1000-2006(2024)02-0247-09
Modelling the potential distribution area of" Populus davidiana" in China based on the Biomod2
GAO Minglong1, TIE Niu2, ZHANG Chen1, LI Fengzi1, WU Yahan1, LUO Qihui1," WANG Zirui1, LIU Lei1,SA Rula1
(1. Forestry College of Inner Mongolia Agricultural University, Hohhot 010019, China; 2. Forestry and Grassland Bureau of Inner Mongolia Autonomous Region, Hohhot 010020, China)
Abstract: 【Objective】 This study aims to" investigate the effects of changes in environmental factors on the distribution of Populus davidiana, and to provide theoretical support for the conservation and development of P. davidiana resources. 【Method】 This study applied Biomod2 to simulate changes in the spatial distribution pattern of P. davidiana in China’s potential distribution areas under three future climatic conditions based on 134 geographical distribution data points of P. davidiana in China, combined with 18 climatic, soil and topographic factors. Then a combinatorial model based on the Biomod2 package was consturcted and identified the main environmental variables affecting the distribution of P. davidiana were identified. 【Result】 The"" current potential distribution areas of P. davidiana in China were mainly located at higher latitudes or higher altitudes on both sides of the 400 mm precipitation line, with a total area of about 1 560 340.9 km2, of which the Greater Khingan Mountains, Changbai Mountains, Taihang Mountains, Qinling Mountains, southern foot of Qilian Mountains, Hengduan Mountains, Yunnan-Guizhou Plateau and other areas are the" highest suitable areas for P. davidiana. Under future climatic conditions, the overall trend of suitable areas for P. davidiana will shrink to southwest China, and the overall trend of suitable areas was decreasing. The ensemble model constructed based on the five optimal single models had better prediction results for suitable areas for P. davidiana compared to the single model, and the area under the receiver operating characteristic curve and true skill statistics were distributed as 0.91 and 0.73, with higher prediction accuracy. 【Conclusion】The spatial distribution pattern of P. davidiana in China was mainly influenced by water and heat conditions, while altitude was also an important factor affecting its distribution. Under future climatic conditions, the area of P. davidiana distribution will gradually decrease based on the degree of climate warming. When planting P. davidiana for timber forests and as an ecological public welfare forest species, planting sites should be selected in areas where habitat suitability will not change significantly in the future, to reduce future losses because of climate change.
Keywords:Populus davidiana; Biomod2; ensemble model; potential distribution area; global warming
全球气候变暖会对物种分布造成严重影响,气候持续变暖将使得温带森林植被向高纬度或高海拔地区迁移。近年来分析气候变化对物种分布范围的影响已经成为生态学的研究热点之一。因此开展气候变化对森林物种潜在分布区影响的研究,对未来我国森林资源开发与环境保护具有重要的理论指导意义。物种分布模型可依据物种实际地理分布数据和相关环境变量对物种潜在分布区进行模拟67〗。目前,主要使用的物种分布模型有广义线性模型(GLM)、推进式回归树模型(GBM)、分类与回归树模型(CTA)、人工神经网络(ANN)、表面分布区分室模型(SRE)、柔性判别分析(FDA)、随机森林(RF)、最大熵值模型(MaxEnt)等。其中,最大熵值模型在酸枣(Ziziphus jujuba var. spinosa)、杉木(Cunninghamia lanceolata)、白杄(Picea meyeri)等大量物种的分布研究中均有应用。然而仅使用一种模型对物种潜在分布区进行预测时,单一模型结果往往稳定性不强、偏差较大,而基于多模型构建的组合模型在准确度上的表现则相对更佳。因此,目前物种分布研究所使用的物种分布模型逐渐由单一模型转向组合模型。
杨柳科(Salicaceae)杨属(Populus )树种广泛分布于北半球,其种植面积和木材产量在所有树种中均位居前列,是世界上重要的绿化树种和用材树种之一。其中山杨(P. davidiana)是我国温带常见的落叶乔木,常作为森林更新的先锋树种,其树干通直、材质优良,木材质地轻软、弹性较好,是造纸、建筑等行业的常用原料,同时山杨在绿化环境、保持水土方面也具有较大作用,生态及经济价值较高。因此,确定我国山杨不同气候背景下的潜在分布区变化,可为我国未来山杨资源的高效利用与保护提供参考。
1" 材料与方法
1.1" 山杨分布数据选择
通过查询2017—2021年的野外实地调查及国家标本馆(NSⅡ)、中国数字标本馆(CVH)、全球生物多样性信息网络数据库(GBIF)等公开文献相关资料,共获得山杨237个分布点数据,剔除其中地理信息标注不明及重复的分布点,最终得到中国境内134个山杨分布点。
1.2" 环境变量筛选及模型构建
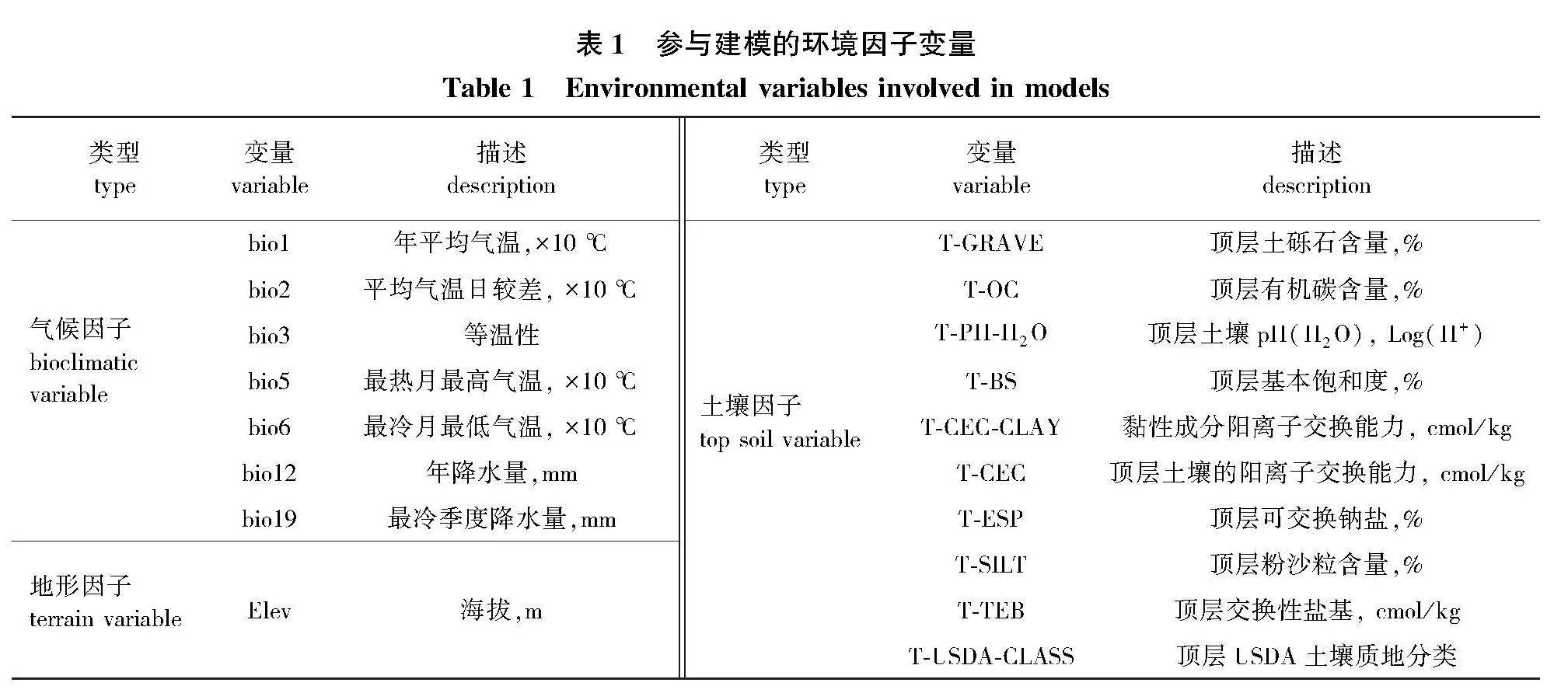
初步选用空间分辨率均为30″的34个环境因子作为建模备选环境因子,其中19个生物气候因子来自世界气候数据库(www.worldclim.org)。以第二代国家(北京)气候中心气候系统模式(BCC-CSM2-MR)作为未来气候系统模式,采用不同温室气体排放浓度和社会发展水平下的SSP126、SSP245和SSP585气候预测结果作为从低到高3个等级的气候情景代表未来全球气候变暖的3种趋势。则本研究中气候情景包括末次冰盛期(LGM)、全新世中期(MH)当前(current)及SSP126-2050s、SSP126-2090s、SSP245-2050s、SSP245-2090s、SSP585-2050s、SSP585-2090s。土壤和地形数据选用世界土壤数据库(www.fao.org.)提供的14个表层土壤和1个地形因子数据。地图数据为自然资源部标准地图服务网站(http:∥www.mnr.gov.cn/)提供的中国标准地图,审图号为GS(2019)1698号。
为避免各环境因子之间过高共线性所导致的模型过度拟合,本研究通过R 4.3.1中usdm软件包对各环境因子进行方差膨胀因子检验(VIF)和Pearson相关性检验,保留其中相关系数小于0.8、VIF值小于10的因子参与最终的建模。最终,获得7个气候因子、10个土壤因子和1个地形因子供建模使用(表1)。
运用Biomod2软件包对山杨的分布区建模,所选用8种模型为: 广义线性模型(GLM)、推进式回归树模型(GBM)、分类与回归树模型(CTA)、人工神经网络(ANN)、表面分布区分室模型(SRE)、柔性判别分析(FDA)、随机森林(RF)、最大熵值模型(MaxEnt)。除MaxEnt模型采用ENMeval程序包对其进行优化后的参数(调控倍频RM为0.5,特征组合为LQ)进行建模外,其余7种模型均采用Biomod2默认模型参数设置。在建模过程中,随机选取山杨134个分布点中的75%作为训练数据集,25%作为测试数据集。此外,为满足Biomod2建模要求并更好地模拟实际分布,本研究随机生成500个伪缺失点参与建模。为避免单次建模所产生的误差,每种模型对上述过程进行15次重复,最终共生成120个建模结果。
参与建模各环境因子的重要性通过Biomod2软件包分析评价得出,各单一模型准确度通过受试者工作特征曲线(ROC)和真实技巧统计值(TSS)进行检验。使用ROC曲线下面积(AUC)评估模型精度时,AUC取值在0.5~1.0之间,0.5代表完全随机的分类;1.0表示完全正确地分类,当AUC值高于0.8表示模型预测性能较好或非常好,低于0.7表示模型预测结果较差。TSS值表示实测样本上的净预测成功率,其取值在0~1之间,当TSS值>0.7时,表示模型预测精度较高;当TSS值<0.5时,则表示模型精度较差。根据单一模型评价结果,在所生成的120个建模结果中选取TSS值和AUC值最高的5个建模结果构建组合模型,根据单模型TSS值评估分数大小分配各单一模型在组合模型中的权重。
1.3" 数据处理
将筛选后的环境数据及山杨分布点数据输入构建的组合模型,运用ArcGIS 10.4.1将模型运行后生成的数据进行可视化,根据模型生成的山杨适宜性分布阈值,将山杨生境适宜度划分为4个等级,依次为:非适生区。根据不同时期山杨适生区和非适生区的地理空间变化,在ArcGIS软件中计算并绘制未来气候变化背景下的山杨未来空间分布格局变化图;使用SDMtoolbox 2.0工具包计算不同气候背景下山杨适生区3个时期的质心位置以及迁移方向,并运用Matlab 2016根据地球曲率计算其质心迁移距离。
2" 结果与分析
2.1" 模型精度评价
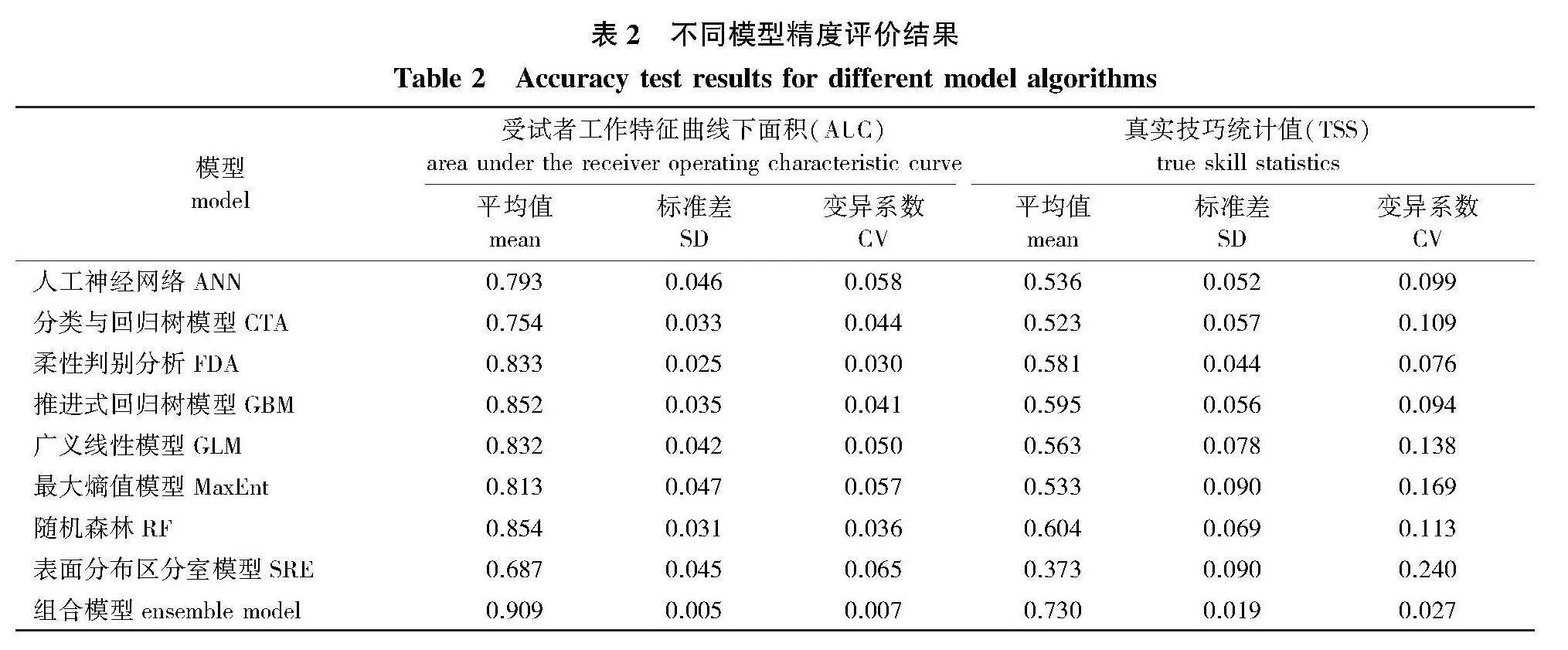
所选8种单一模型的预测精度如表2所示,随机森林(RF)模型为最优模型,其AUC值和TSS值均为最高,多次建模稳定性也相对较好;表面分布区分室模型(SRE)预测精度最差,AUC值和TSS值仅为0.687、0.373,无法对适生区分布进行正确模拟。除SRE模型外,其余模型AUC值和TSS值的均值均大于0.70和0.50,预测准确率相对较好。与单一模型相比,基于5个最优单一模型(RF、GBM、GLM、MaxEnt、FDA)构建的组合模型预测精度有较大幅度提升,其AUC值为0.909、TSS值为0.730,对山杨适生区的预测达到了较好的水平。因此,本研究后续内容均基于组合模型运行结果进行分析。
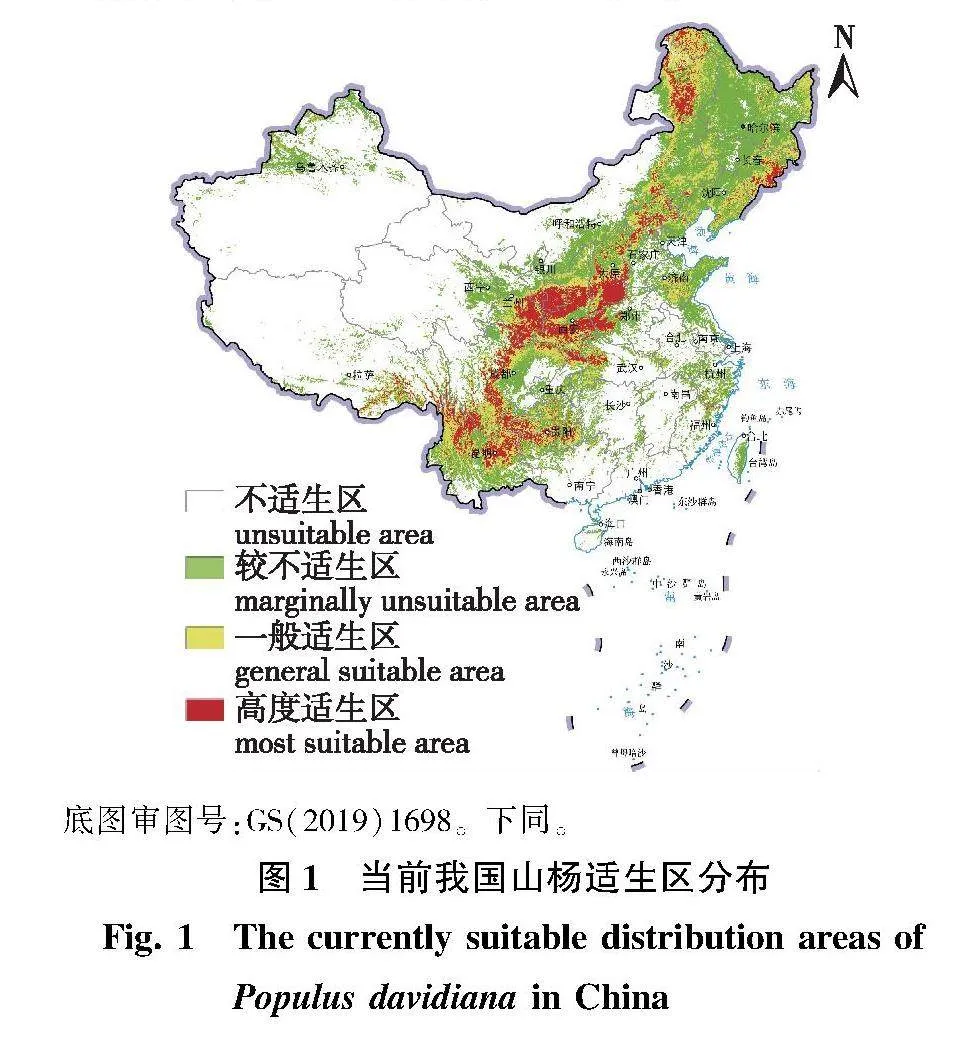
2.2" 当前和过去时期我国山杨地理分布
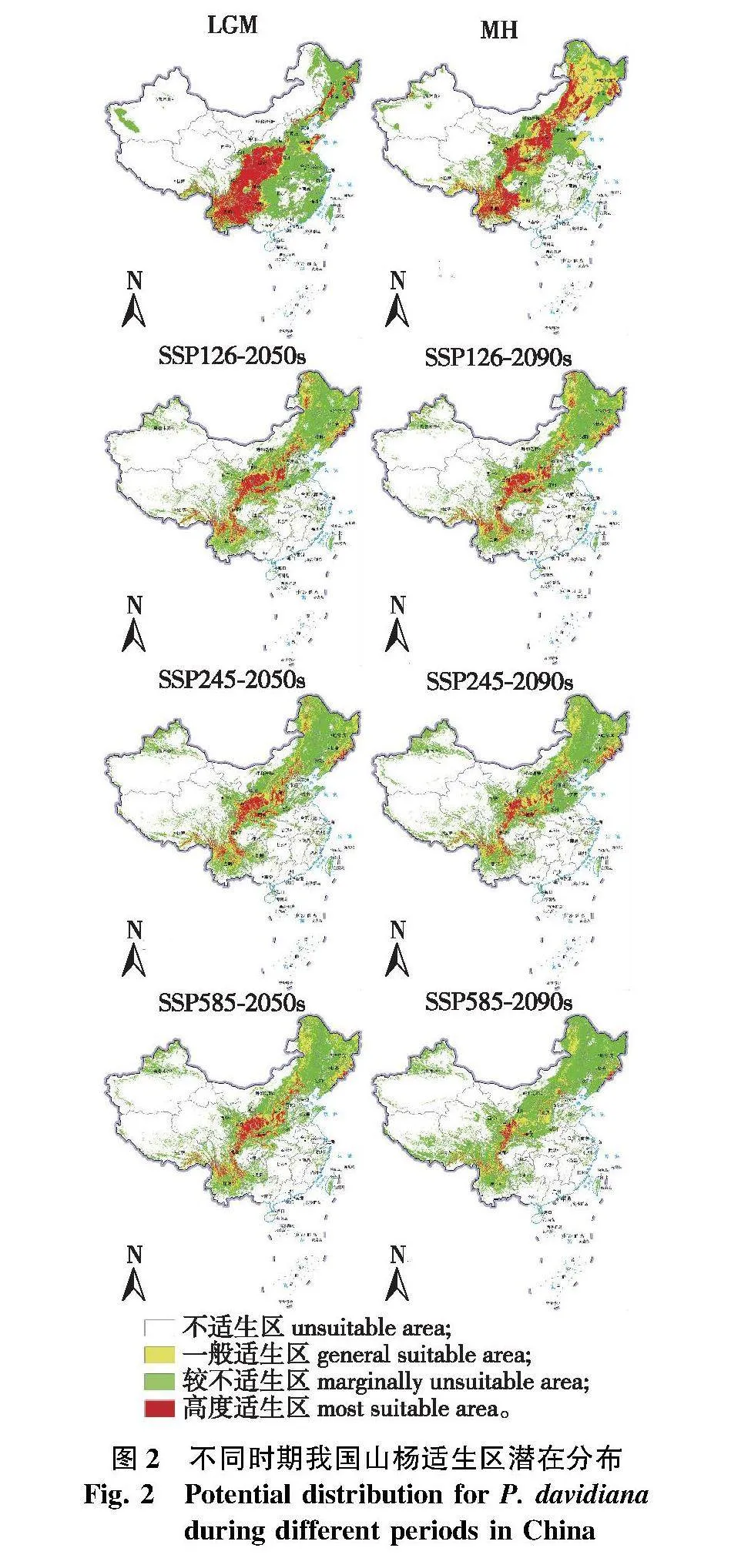
根据组合模型预测结果(图1),当前山杨适生区(一般适生区与高度适生区之和)大体在400 mm等降水线两侧分布,主要分布于内蒙古、黑龙江、吉林、辽宁、河北、北京、陕西、山西、河南、宁夏、甘肃、山东、四川、重庆、湖北、贵州、云南及西藏等地,在新疆、青海、安徽、江西、湖南、广西等地的较高海拔地区有少量分布;适生区面积为1 560 340.9 km2,其中高度适生区面积为553 489.5 km2,一般适生区面积为1 006 851.4 km2。山杨高度适生区主要分布在大兴安岭、长白山、太行山、秦岭、祁连山南麓、横断山、云贵高原等地区。
在末次冰盛期,山杨总适生区面积约为1 317 834 km2,分布格局与当前时期相比差异较大,适生区主要位于我国云南、贵州、四川、重庆、陕西等省市,在山东半岛、长白山地区和东北平原局部地区存在小面积分布(图2)。
在全新世中期,山杨总适生区发生明显扩张,面积约为2 327 603 km2,相较末次冰盛期扩大176.6%,分布区域与当前山杨分布情况较为相似,此时西南地区适生区开始缩小并呈现出破碎化趋势,东北地区山杨适生区相较当前气候条件分布面积更大。在过去2个时期中,长白山地区和横断山脉部分地区均为山杨主要分布区,与当前山杨适生区相比变化较小。
2.3" 未来山杨适生区预测及质心迁移路线
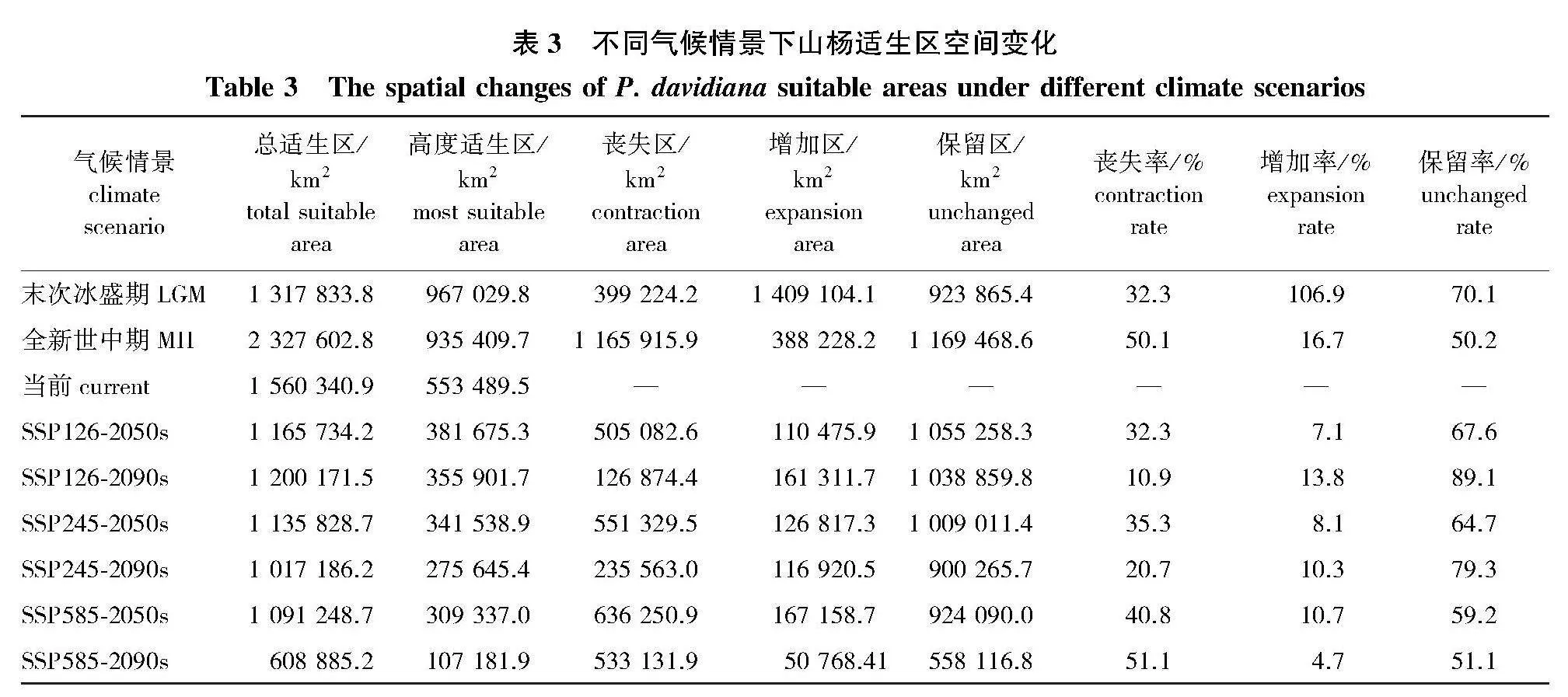
在未来时期,除SSP126气候情境下山杨总适生区面积为减少后小幅增加,其余气候情景下2050s和2090s山杨总适生区面积与上一时期相比均有较大的缩减(表3和图2)。在SSP126气候情景下,山杨总适生区面积变化最小,2050s总适生区面积相较当前缩小25.3%,约合面积为394 607 km2;2090s总适生区面积相较2050s小幅度增加2.9%,增加面积为34 437 km2。在SSP585气候情景下,2050s总适生区面积相较当前总适生区缩小27.2%,缩小面积约为424 512 km2;至2090s总适生区面积再次缩小10.2%,面积约为118 642 km2。在SSP245气候情景下,2050s总适生区面积相较当前总适生区缩小30.1%,约合面积为469 092 km2;2090s总适生区面积缩小44.2%,约合面积为482 363 km2。由此可见,无论是在2050s还是2090s,SSP585气候情景下山杨总适生区面积变化幅度在3种气候情境中均为最大。
通过未来2个时期适生区的变化可知,不同气候情景对山杨适生区的影响不尽相同,在SSP585气候情景下山杨适生区对气候变化最为敏感(图2、图3和表3)。在SSP126情景下,山杨高度适生区面积的变化幅度最小,2090s山杨总适生区面积出现小幅增加,但此时高度适生区面积相较2050s减少6.7%,面积约为25 773.6 km2,这说明2090s适生区新增面积以一般适生区面积为主。在SSP585情景下,2050s山杨高度适生区面积的损失最大,同时新增加适生区面积最大,相较当前其高度适生区面积减少44.1%,面积约为244 152.5 km2;到2090s高度适生区面积缩减比例最大,相较2050s减少65.3%,面积约为202 155.1 km2,同时新增适生区面积最小,仅为50 768.41 km2。在SSP245情景下,山杨高度适生区的变化幅度介于SSP126和SSP585气候情景之间,2050s高度适生区面积相较当前减少38.2%,2090s高度适生区面积相较当前减少19.3%。
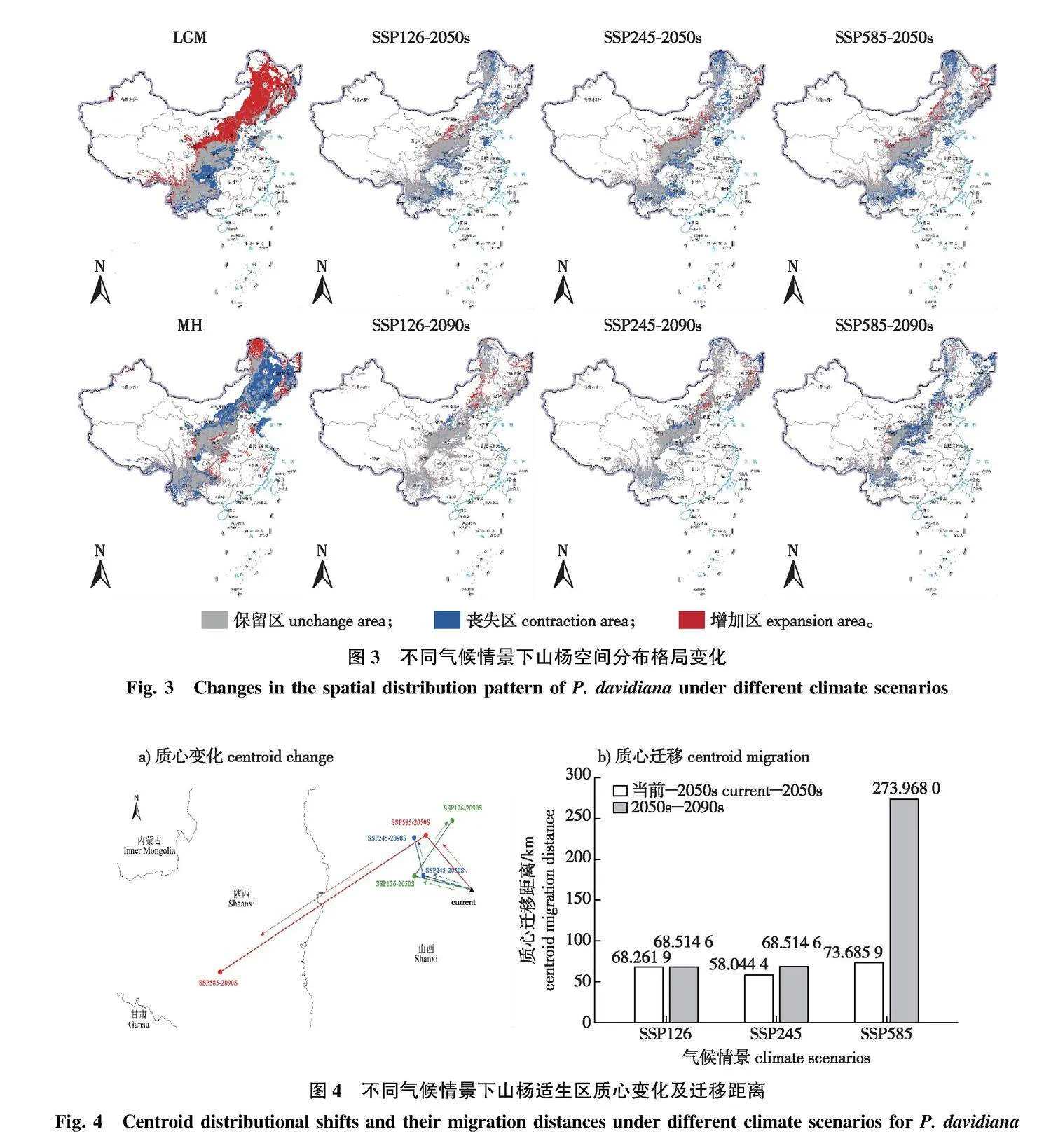
在空间格局方面,山杨适生区的质心除SSP585-2090s气候情境下向西南大幅迁移外,其余时期均为小幅向北迁移(图4)。当前时期山杨适生区的质心位于山西省晋中市太谷区(112°40′17″E, 37°21′20″N)。当气候情景为SSP585-2090s时,山杨适生区质心迁移幅度最大,迁移距离可达273.9 km,此时山杨适生区质心位于陕西省延安市安塞区(109°17′39″E, 36°49′4″N);当气候情景为SSP126-2090s时,山杨适生区质心位置为各时期各气候背景下的最北端,位于山西省太原市尖草坪区(109°38′24″E, 40°38′24″N),距离为68.3 km。在未来全球增温增湿的气候变化情景下,中国山杨适生区的质心发生小幅度向北迁移,但随着气候进一步变化,山杨适生区质心转为向西南迁移。
2.4" 影响山杨分布的环境因子分析
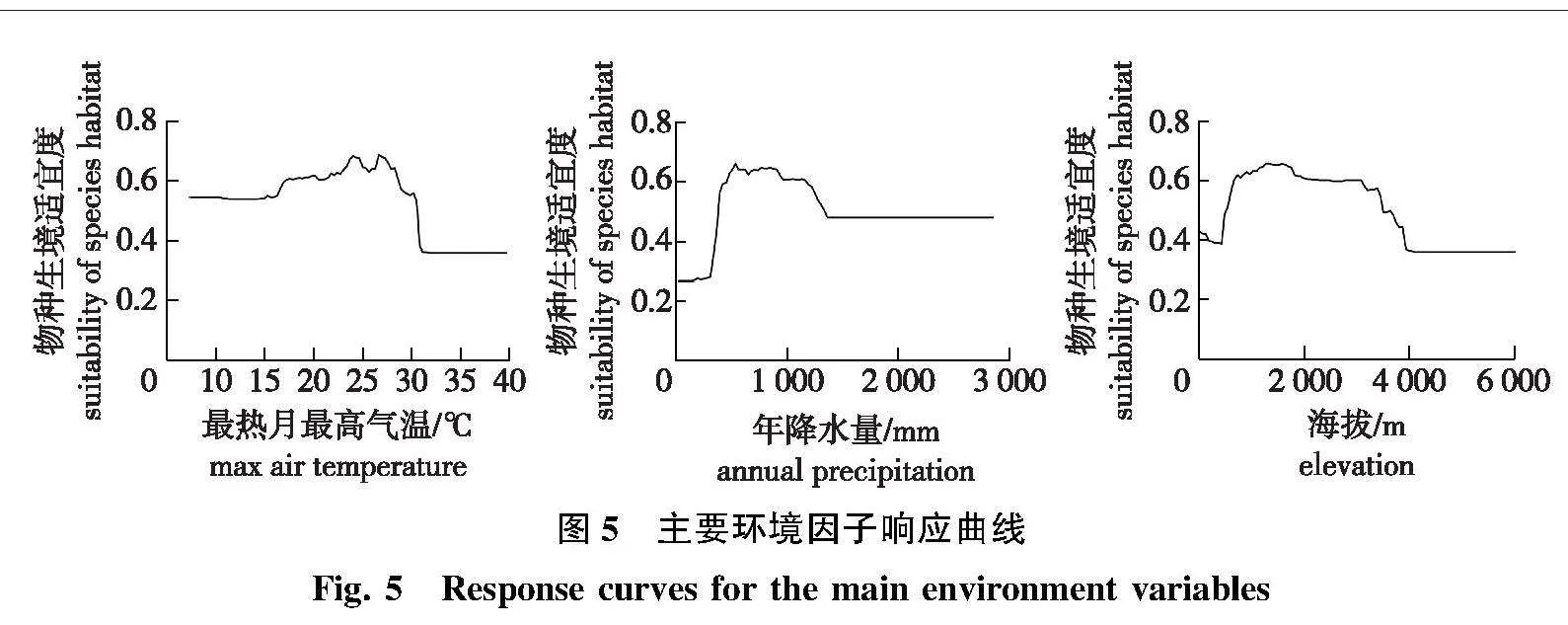
根据Biomod2软件包的运行结果,所选环境因子中对山杨潜在地理分布影响最大的5个环境因子及其重要度分别为:最热月最高气温(bio5,0.121)、年降水量(bio12,0.103)、海拔(Elev,0.051)、平均气温日较差(bio2,0.031)、最冷季度降水量(bio19,0.028)。其中,最热月最高气温(bio5)、年降水量(bio12)和海拔(Elev)的重要性占比高达63.4%。由此可知,影响山杨分布的主要环境因子为最热月最高气温、年降水量和海拔(图5)。
由主要环境因子响应曲线可知,山杨的物种生境适宜度受最热月最高气温、年降雨量和海拔的显著影响。山杨的物种生境适宜度在最热月最高气温为30 ℃左右时明显下降,山杨的物种生境适宜度与年降雨量和海拔响应曲线均为先增加后降低的倒“V”形。这表明山杨更适宜分布于最热月最高气温在30 ℃以下、年降水500~1 250 mm且海拔在700~3 500 m之间的地区。
2.5" 山杨未来分布区生态特征变化
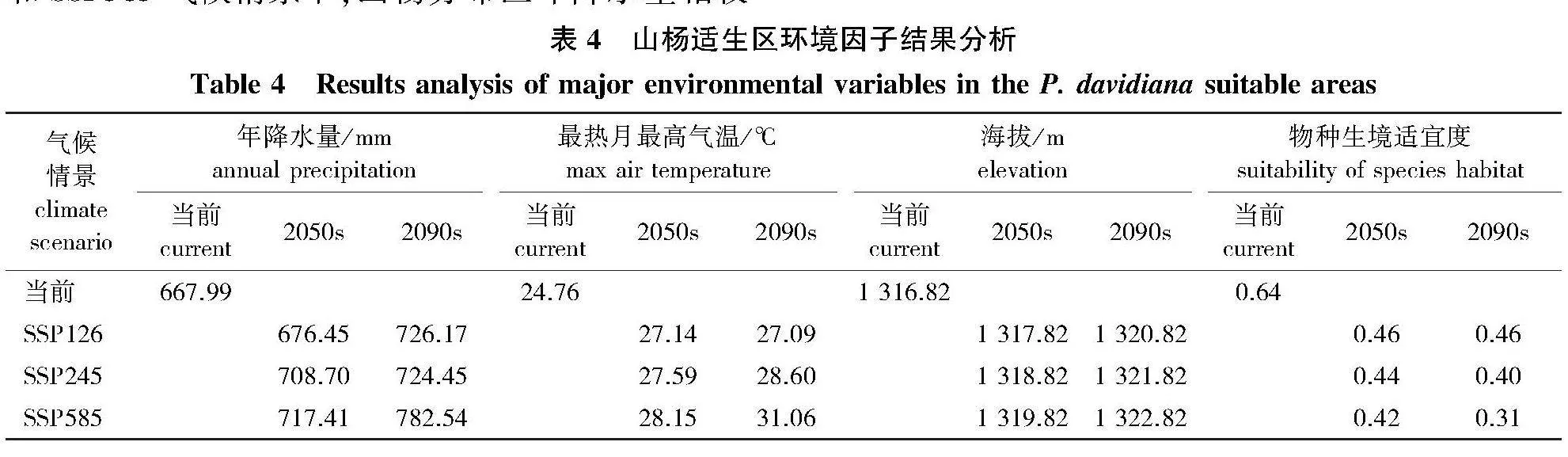
主要环境因子与山杨生境适宜度的关系见表4。在SSP126气候情景下,2050s和2090s山杨现实分布点生境适宜度均比当前低0.18;在SSP245气候情景下,2050s和2090s生境适宜度分别比当前低0.20和0.24;在SSP585气候情景下,山杨生境适宜度在2090s下降较大,相较当前降低51.56%,仅为0.31,这表明在该气候条件下当前的部分山杨分布点已不再适合山杨生长。
山杨分布区年降水量与山杨生境适宜度的变化趋势大体相反。在2050s时期SSP126、SSP245和SSP585气候情景下,山杨分布区年降水量相较当前时期分别增加8.46、40.71和49.42 mm;2090s与当前相比,SSP126、SSP245和SSP585情景的年降水量分别增加58.18、56.46和114.55 mm。
山杨分布区的最热月最高气温在SSP126、SSP245和SSP585气候条件下均逐渐升高,与年降水量类似,最热月最高气温也与山杨生境适宜度变化趋势相反。在2050s,SSP126、SSP245和SSP585气候条件下最热月最高气温相较当前分别增加2.38、2.83和3.39" ℃;2090s与当前相比,SSP126、SSP245和SSP585气候条件下最热月最高气温分别增加2.33、3.84和6.30" ℃。
3" 讨" 论
山杨在我国分布广泛,黑龙江、内蒙古、吉林、华北、西北、华中及西南地区均有分布。通过分析各单一模型预测结果和山杨实际分布区域的对比结果,以及各单一模型的AUC值和TSS值发现,随机森林(RF)、优化后的最大熵值模型(MaxEnt)、广义线性模型(GLM)及推进式回归树模型(GBM)对山杨适生区均取得较好的模拟结果。选取5个最优单一模型构建的组合模型在山杨分布区预测精度上相较8类单一模型均有较大提升,AUC值和TSS值相较单一模型中表现最好的随机森林模型(RF)分别提升6.4%和20.9%,同时还解决了MaxEnt等模型对山杨适生区细节刻画精度不高的问题。这表明相较于单一模型对山杨的分布区预测,组合模型的预测结果更为准确。
山杨分布受各种环境条件的制约,其中最热月最高气温、年降水量和海拔是影响山杨分布的主要环境因子。本研究发现,山杨分布区的年降水量与山杨生境适宜度呈先增加后减少的倒“V” 形关系,山杨适生区在南北走向上大体沿400 mm等降水线两侧分布,表明过高或过低的降水量均不利于山杨生长分布。同时,平均最热月最高温与山杨生境适宜度呈负相关关系,平均最热月最高气温高于30 ℃时山杨生境适宜度出现大幅下降,这与山杨作为一种耐寒冷、耐干旱、强阳性树种的生物学特性相符。而山杨适生区海拔从东北至西南逐渐增加,平均海拔达1 316.82 m,高度适生区主要分布于高海拔山地且呈碎片状分布,这又从另一方面证明了山杨对寒冷、干旱有较高的耐受性,同时也印证了前人对山杨分布区海拔南高北低且西南地区的山杨多分布于高山地区的调查结果。此外,本研究选用的10个土壤因子对山杨生长和分布的影响均相对较小,土壤因子重要度之和占比仅为10.4%,说明山杨对土壤条件要求不高,具有较宽的适应范围。本研究推测正是因为山杨耐寒耐旱耐贫瘠的特殊生物学特性,可使其在其他阔叶树种难以分布的高寒、高海拔地区生长分布,也可以从侧面解释山杨在我国众多地区出现大范围分布的现象。
本研究选取的2个历史时期中,末次冰盛期气候寒冷干旱,全新世中期气候与当前气候条件最为相似。相关研究表明,在末次冰盛期,温带树种在高纬度地区分布范围发生收缩;在全新世中期,温带树种适宜生境有较大程度的扩张。山杨过去时期适生区变化趋势与该结论基本一致。在末次冰盛期,山杨适生区整体面积较小且连续分布区主要分布在西南较为温暖地区。在全新世中期,山杨适生区发生大面积的扩张,逐渐形成与当前类似的地理分布格局;但全新世中期暖温带潮湿气候区分布更广,导致当前气候条件下的干旱地区也适宜山杨生长分布,山杨适生区面积较当前时期更为广阔。
在本研究模拟的3种未来气候情境中,山杨适生区面积随气候增温增湿呈现出先减小后小幅增加再转为大幅减小的波动变化趋势。在SSP126气候情景下,2050s山杨适生区面积相较当前适生区面积有较大幅度的减少,2090s相较2050s有小幅度增加。而在全球增温增湿更为剧烈的SSP245和SSP585气候情境下,山杨适生区面积均随气候变化幅度的增加而减小。这种变化特征表明山杨适生区的面积变化对气候变暖的响应存在一个区间,即当气候变暖程度在此区间内时增温增湿有利于山杨分布区扩张,反之不利。本研究由于未使用连续气候变化参数,具有一定的局限性,只能推测出该区间的存在,但此区间具体数值范围仍有待进一步研究。
在未来全球气候变暖趋势下,部分物种通过其生物学特性的变化来适应气候变化,而有些物种则选择向高纬度或高海拔地区迁移来规避气候变化造成的影响。本研究中,山杨在未来2个时期3种气候情境下均明显出现了向高海拔和高纬度地区迁移的趋势。SSP126和SSP245气候情境下,山杨适生区质心在2050s和2090s均不同程度地向北迁移。然而,在SSP585气候情境下,2090s山杨适生区的质心表现出向北迁移后转为向南迁移的趋势。本研究通过对比横向各时期山杨适生区的变化发现:在SSP585极端气候条件下我国北方低海拔地区山杨适生区在2090s大面积丧失,而西南地区山杨在垂直方向上具有更大的迁移空间,适生区面积损失较小,南北地区山杨适生区的不同变化造成了SSP585-2090s气候情景下特殊的质心迁移路径。由此推测,如果未来全球变暖程度相较SSP585气候情景进一步加剧,我国西南高山地区则可能成为山杨的气候避难所。
本研究通过Biomod2软件包构建组合模型对不同气候情景下山杨适生区进行预测,相较于传统单一模型,组合模型预测准确度有较大提高,预测结果与山杨实际分布基本一致。中国山杨当前适生区主要分布于400 mm等降水线两侧较高纬度或较高海拔地区,且南方高度适生区相较北方更为破碎。分布区生态特征变化表明,环境因子中的最热月最高温、年降水量和海拔对山杨生长分布影响最大;在全球变暖的背景下,全国各地山杨适生区整体上呈收缩趋势,并向高海拔或高纬度地区迁移。在极端气候情景下,山杨在我国北方低海拔地区的适生区将大面积收缩,横断山脉、四川盆地与青藏高原交界等西南较高海拔地区将成为山杨主要分布区。以山杨作为用材林和生态公益林树种进行造林时,造林地点应选择未来生境适宜度变化不大的地区,如长白山、祁连山南麓及横断山脉等地区,以降低未来由于气候变化造成的损失。
参考文献(reference):
[1]曾建平,代峰.气候伦理是否可能.中国人民大学学报,2011,25(3):90-96.ZENG J P,DAI F.The possibility of a climatic ethics.J Renmin Univ China,2011,25(3):90-96.
[2]BEZENG B S,MORALES-CASTILLA I,VAN DER BANK M,et al.Climate change may reduce the spread of non-native species.Ecosphere,2017,8(3):e01694.DOI: 10.1002/ecs2.1694.
[3]BELLARD C,BERTELSMEIER C,LEADLEY P,et al.Impacts of climate change on the future of biodiversity.Ecol Lett,2012,15(4):365-377.DOI: 10.1111/j.1461-0248.2011.01736.x.
[4]VILLN-PERZ S,HEIKKINEN J,SALEMAA M,et al.Global warming will affect the maximum potential abundance of boreal plant species.Ecography,2020,43(6):801-811.DOI: 10.1111/ecog.04720.
[5]ELITH J,GRAHAM C H,ANDERSON R P,et al.Novel methods improve prediction of species distributions from occurrence data.Ecography,2006,29(2):129-151.DOI: 10.1111/j.2006.0906-7590.04596.x.
[6]刘晓彤,袁泉,倪健.中国植物分布模拟研究现状.植物生态学报,2019,43(4):273-283.LIU X T,YUAN Q,NI J.Research advances in modelling plant species distribution in China.Chin J Plant Ecol,2019,43(4):273-283.DOI: 10.17521/cjpe.2018.0237.
[7]ZHAO G H,CUI X Y,SUN J J,et al.Analysis of the distribution pattern of Chinese Ziziphus jujuba under climate change based on optimized biomod2 and MaxEnt models.Ecol Indic,2021,132:108256.DOI: 10.1016/j.ecolind.2021.108256.
[8]TANG J H,LI J H,LU H,et al.Potential distribution of an invasive pest,Euplatypus parallelus,in China as predicted by Maxent.Pest Manag Sci,2019,75(6):1630-1637.DOI: 10.1002/ps.5280.
9〗赵光华,崔馨月,王智,等.气候变化背景下我国酸枣潜在适生区预测.林业科学,2021,57(6):158-168.ZHAO G H,CUI X Y,WANG Z,et al.Prediction of potential distribution of Ziziphus jujuba var.spinosa in China under context of climate change.Sci Silvae Sin,2021,57(6):158-168.DOI: 10.11707/j.1001-7488.20210618.
LI Y C,LI M Y,LI C,et al.Optimized maxent model predictions of climate change impacts on the suitable distribution of Cunninghamia lanceolata in China.Forests,2020,11(3):302.DOI: 10.3390/f11030302.
吴晓萌,叶冬梅,白玉娥,等.基于MaxEnt模型的中国白杄分布格局及未来变化.西北植物学报,2022,42(1):162-172.WU X M,YE D M,BAI Y E,et al.Distribution pattern and future change of Picea meyeri in China based on MaxEnt model.Acta Bot Boreali Occidentalia Sin,2022,42(1):162-172.DOI: 10.7606/j.issn.1000-4025.2022.01.0162.
GONG Y F,HU X K,HAO Y W,et al.Projecting the proliferation risk of Oncomelania hupensis in China driven by SSPs:a multi-scenario comparison and integrated modeling study.Adv Clim Change Res,2022,13(2):258-265.DOI: 10.1016/j.accre.2022.02.004.http://dx.doi.org/10.1016/j.accre.2022.02.004
HAO T X,ELITH J,GUILLERA-ARROITA G,et al.A review of evidence about use and performance of species distribution modelling ensembles like BIOMOD.Divers Distrib,2019,25(5):839-852.DOI: 10.1111/ddi.12892.
卫尊征,郭丽琴,张金凤,等.利用trnL-F序列分析杨属树种的系统发育关系.北京林业大学学报,2010,32(2):27-33.WEI Z Z,GUO L Q,ZHANG J F,et al.Phylogenetic relationship of Populus by trnL-F sequence analysis.J Beijing For Univ,2010,32(2):27-33.DOI: 10.13332/j.1000-1522.2010.02.031.
中国科学院中国植物志编辑委员会.中国植物志-第二十卷,第二分册.北京:科学出版社,1984.Delectis Florae Reipublicae Popularis Sinicae,Agendae Academiae Sinicae Edita.Flora reipublicae popularis sinicae tomus 20(2).Beijing:Science Press,1984.
翁宇威,蔡闻佳,王灿.共享社会经济路径(SSPs)的应用与展望.气候变化研究进展,2020,16(2):215-222.WENG Y W,CAI W J,WANG C.The application and future directions of the Shared Socioeconomic Pathways(SSPs).Clim Change Res,2020,16(2):215-222.DOI: 10.12006/j.issn.1673-1719.2019.078.
YANG X Q,KUSHWAHA S P S,SARAN S,et al.MaxEnt modeling for predicting the potential distribution of medicinal plant,Justicia adhatoda L.in Lesser Himalayan foothills.Ecol Eng,2013,51:83-87.DOI: 10.1016/j.ecoleng.2012.12.004.
张天蛟,刘刚.提高生态位模型时间转移能力的方法研究.中国农业大学学报,2017,22(2):98-105.ZHANG T J,LIU G.Study of methods to improve the temporal transferability of niche model.J China Agric Univ,2017,22(2):98-105.DOI: 10.11841/j.issn.1007-4333.2017.02.12.
DORMANN C F,ELITH J,BACHER S,et al.Collinearity:a review of methods to deal with it and a simulation study evaluating their performance.Ecography,2013,36(1):27-46.DOI: 10.1111/j.1600-0587.2012.07348.x.
ALLOUCHE O,TSOAR A,KADMON R.Assessing the accuracy of species distribution models:prevalence,Kappa and the true skill statistic (TSS).J Appl Ecol,2006,43(6):1223-1232.DOI: 10.1111/j.1365-2664.2006.01214.x.
吴艺楠,马育军,刘文玲,等.基于BIOMOD的青海湖流域高原鼠兔分布模拟.动物学杂志,2017,52(3):390-402.WU Y N,MA Y J,LIU W L,et al.Modelling the distribution of plateau pika(Ochotona curzoniae) in Qinghai Lake basin using BIOMOD.Chin J Zool,2017,52(3):390-402.DOI: 10.13859/j.cjz.201703004.
郭恺琦,姜小龙,徐刚标.薄片青冈潜在适生区及气候变化对其分布的影响.生态学杂志,2021,40(8):2563-2574.GUO K Q,JIANG X L,XU G B.Potential suitable distribution area of Quercus lamellosa and the influence of climate change.Chin J Ecol,2021,40(8):2563-2574.DOI: 10.13292/j.1000-4890.202108.023.
ZHAN P,WANG F Y,XIA P G,et al.Assessment of suitable cultivation region for Panax notoginseng under different climatic conditions using MaxEnt model and high-performance liquid chromatography in China.Ind Crops Prod,2022,176:114416.DOI: 10.1016/j.indcrop.2021.114416.
李易.中国山杨群体历史动态初步研究.北京:中国林业科学研究院,2020.LI Y.A preliminary study on demographic history of Populus davidiana.Beijing:Chinese Academy of Forestry,2020.
郭彦龙,赵泽芳,乔慧捷,等.物种分布模型面临的挑战与发展趋势.地球科学进展,2020,35(12):1292-1305.GUO Y L,ZHAO Z F,QIAO H J,et al.Challenges and development trend of species distribution model.Adv Earth Sci,2020,35(12):1292-1305.DOI: 10.11867/j.issn.1001-8166.2020.110.
袁喆,严登华,杨志勇,等.1961—2010年中国400 mm和800 mm等雨量线时空变化.水科学进展,2014,25(4):494-502.YUAN Z,YAN D H,YANG Z Y,et al.Research on temporal and spatial change of 400 mm and 800 mm rainfall contours of China in 1961 to 2000.Adv Water Sci,2014,25(4):494-502.DOI: 10.14042/j.cnki.32.1309.2014.04.002.
贺敏,魏江生,石亮,等.大兴安岭南段山杨径向生长和死亡对区域气候变化的响应.生态学杂志,2018,37(11):3237-3244.HE M,WEI J S,SHI L,et al.The response of radial growth and death of Populus davidiana to regional climate change in southern Greater Xing’an Mountains.Chin J Ecol,2018,37(11):3237-3244.DOI: 10.13292/j.1000-4890.201811.026.
潘春芳,赵秀海,夏富才,等.长白山山杨种群的性比格局及其空间分布.生态学报,2011,31(2):297-305.PAN C F,ZHAO X H,XIA F C,et al.Sex ratio and spatial pattern in Populus davidiana in Changbai Mountain.Acta Ecol Sin,2011,31(2):297-305.
DIAS J M A,BOSKI T,RODRIGUES A,et al.Coast line evolution in Portugal since the Last Glacial Maximum until present: a synthesis.Mar Geol,2000,170(1/2):177-186.DOI: 10.1016/S0025-3227(00)00073-6.
TARASOV P,BEZRUKOVA E,KARABANOV E,et al.Vegetation and climate dynamics during the Holocene and Eemian interglacials derived from Lake Baikal pollen records.Palaeogeogr Palaeoclimatol Palaeoecol,2007,252(3/4):440-457.DOI: 10.1016/j.palaeo.2007.05.002.
白伟宁,张大勇.植物亲缘地理学的研究现状与发展趋势.生命科学,2014,26(2):125-137.BAI W N,ZHANG D Y.Current status and future directions in plant phylogeography.Chin Bull Life Sci,2014,26(2):125-137.DOI: 10.13376/j.cbls/2014020.
HOU Z,LI A,ZHANG J G.Genetic architecture,demographic history,and genomic differentiation of Populus davidiana revealed by whole-genome resequencing.Evol Appl,2020,13(10):2582-2596.DOI: 10.1111/eva.13046.
CHEN I C,HILL J K,OHLEMLLER R,et al.Rapid range shifts of species associated with high levels of climate warming.Science,2011,333(6045):1024-1026.DOI: 10.1126/science.1206432.
SUON S,LI Y,POM L,et al.Spatiotemporal analysis of soil moisture drought over China during 2008-2016.J Water Resour Prot,11,700-712.DOI: 10.4236/jwarp.2019.116041.
胡忠俊,张镱锂,刘林山,等.生物避难所及其识别方法评述.生态学杂志,2013,32(12):3397-3406.HU Z J,ZHANG Y L,LIU L S,et al.Refugia and their identification methods:a review.Chin J Ecol,2013,32(12):3397-3406.DOI: 10.13292/j.1000-4890.2013.0518.
(责任编辑" 郑琰燚)
猜你喜欢
南京林业大学学报(自然科学版)(2024年5期)2024-10-09 00:00:00
森林工程(2021年2期)2021-04-14 03:58:24
绿色科技(2019年19期)2019-11-26 11:57:33
科学与财富(2017年10期)2017-05-09 01:34:32
现代农业科技(2017年1期)2017-03-06 12:56:30
商(2016年24期)2016-07-20 11:14:55
商(2016年9期)2016-04-15 08:57:25
经济与管理(2015年2期)2015-03-17 21:34:17
电影文学(2014年4期)2014-02-26 05:04:02
黑龙江生态工程职业学院学报(2013年4期)2013-11-20 11:31:30
