
高粱SBP基因家族鉴定及干旱胁迫下的表达模式分析
2024-06-11 08:04姜昱雯陈满静赵应任艳周棱波沈佳奇邵明波
山东农业科学 2024年3期
姜昱雯 陈满静 赵应 任艳 周棱波 沈佳奇 邵明波

摘要:在高等植物中,SQUAMOSA Promocer Binding Procein-Box(SBP)普遍参与植物的生长、开花调控及细胞程序性死亡等过程。本研究对高粱(Sorghum bicolor L.)SBP转录因子家族进行鉴定并进行生物信息学分析,同时利用qRT-PCR对高粱SBP基因在PEG-6000模拟干旱胁迫下的表达进行分析,以验证其功能。结果表明,从高粱中共鉴定出19个SBP成员,除第8条染色体外,其他9条染色体上均分布有SBP基因:绝大部分SBP蛋白在亚细胞水平定位于细胞核,且均为亲水性蛋白质。系统发育分析结果表明,与水稻类似,这19个高粱SBP基因也可分为3个亚类,分别含有7、5、7个SBP基因:相比于拟南芥,高粱SBP与水稻、玉米的SBP关系更近。启动子顺式作用元件预测结果显示,高粱SBP可能参与了脱落酸、茉莉酸甲酯信号通路,部分成员可能与植物的低温响应、昼夜节律调控有关。qRT-PCR分析结果显示,高粱SBP基因的表达具有组织特异性,有17个基因受干旱胁迫诱导上调表达,且随胁迫时间的延长存在差异化表达模式。本研究结果可为后续研究高粱SBP基因的功能提供理论依据。
关键词:高粱;SBP基因家族;生物信息学分析;基因表达;干旱胁迫
中图分类号:S514:Q78 文献标识号:A 文章编号:1001-4942(2024)03-0001-10
转录因子是生物体中分子调控网络的重要组成部分,控制着植物生长发育、胁迫响应等一系列生物学过程,不同的转录因子家族结合的下游基因启动子基序存在显著差别。SQUAMOSA romoter Binding Protein-Box(SBP)是一类植物特异性的转录因子家族,因其成员含有高度保守的DNA结合结构域SBP而得名。SBP保守结构域由约76个氨基酸残基组成,参与下游基因启动子DNA的GTAC核心基序结合及细胞核定位,并有两个典型的锌指结构。SBP家族成员于1996年首次在金鱼草(Antirrhinum majus)中鉴定获得,即AmSBP1与AmSBP2,且研究发现这两个SBP成员可以直接结合到花分生相关基因SQUA-MOSA的启动子上调控基因表达。其后在多个物种中鉴定到SBP家族成员,如蓝莓(Vacciniumspp.)、甜根子草(Saccharum spontaneum L.)、黑胡椒(Piper nigrum L.)、拟南芥(Arabidopsis thali-ana)及水稻(Oryza sativa)等,其功能涉及到植物信号转导、防御响应、花和果实发育及相变过程等。如在模式植物拟南芥中共鉴定到16个SBP家族成员,其中,SPL3、SPA和SPL5是开花调控基因LEAFY、FRUITFUL和APETALA1的上游调控子,三者功能冗余,共同調控拟南芥的花发育过程:SPL9和SPL15是植物间隔期和分枝的调控子。在黑胡椒中共鉴定到34个SBP成员,其中17个成员上发现有MiR156的结合位点。水稻中共鉴定到19个SBP成员,其中SPL14控制着水稻分蘖,SPL16可以调控水稻籽粒性状和质量。
高粱(Sorghum bicolor L.)是禾本科高粱属一年生草本植物,是世界第五大禾谷类作物,同时也是极为重要的旱粮作物,被广泛应用于酿酒、食用、饲料、制糖及淀粉制造等方面,具有很高的应用价值。2009年高粱全基因组测序完成,推动了其生长发育调控关键基因家族鉴定工作的开展。另外,全球气候的极端变化也使得生物基因家族的鉴定及功能研究变得尤为重要。高粱具有较强的抗旱性,挖掘其抗旱基因对加快其抗旱育种进程、提高产量具有重要意义。本研究对高粱SBP基因家族进行鉴定,并基于基本理化性质分析、系统发育分析、蛋白一级结构及启动子基序预测等对其进行生物信息学分析,同时利用qRT-PCR分析其在高粱不同组织中及在干旱胁迫不同时间下的表达模式,以期为后续深入研究SBP转录因子家族在高粱生长发育过程中的功能并筛选出优良基因奠定基础。
1 材料与方法
1.1 样品处理
受试高粱种子购白红缨子农业科技发展有限公司。2023年10月将种子播于营养丰富的土壤基质中进行光照恒温培养,温度为23℃。待幼苗长出4-5片叶时进行干旱胁迫处理,即将幼苗根部直接浸在10%的PEG-6000溶液中,分别于处理0、4、12、18、24、36h采集样品,并置于液氮中速冻后保存于-80℃冰箱。
1.2 数据来源
拟南芥的SBP基因下载于TAIR(https://ww.arabidopsis.org/),水稻和高粱的SBP家族成员从PlantTFDB(http://planttfdb.cbi.pku.edu.cn/)网站获得。高粱基因组、mRNA、蛋白组及相应的GFF文件来自于Phytozome(http://phyto-zome.jgi.doe.gov/pz/portal.html)数据库。
1.3 试验方法
1.3.1 高粱SBP家族成员鉴定
通过NCBI(ht-tps://www.ncbi.nlm.nih.gov/)的Blast功能,利用拟南芥16个SBP成员的氨基酸序列在高粱蛋白组中进行序列预测(E value≤10-5),获得SBP候选蛋白序列,再通过NCBI的保守结构域分析功能及pfam(http://pfam.xfam.org/)网站检测序列中是否存在SBP结构域。通过TBtools软件提取高粱SBP成员的CDS、启动子序列,方便后续分析。
1.3.2 SBP家族系统发育分析
获得拟南芥、水稻、玉米及高粱的SBP氨基酸序列后,利用MEGA软件使用邻接法(Neighbor-Joining)构建系统发育树,Bootstrap设置为1000,构建好的系统发育树使用Figtree软件进行优化。另用高粱的19个SBP家族成员构建系统发育树,Bootstrap设置为1000。
1.3.3 高粱.SBP基因染色体定位
使用TBtools工具箱中的Gene Location Visualize from GFT/CFF工具,按照操作指示,分别导入高粱基因组的GFF文件和.SBP基因的ID信息,获得染色体定位图片,并用Photoshop软件进行美化调整。
1.3.4 高粱SBP成员的理化性质分析
在Prot-Param(http://us.expasy.0rg/tools/protparam.html)网站上对高粱SBP成员的氨基酸序列长度、等电点、带负电荷残基数、带正电荷残基数、不稳定指数、脂肪族氨基酸指数和平均疏水性进行预测,并利用PSORT(http://psort.hgc.jp/)对其进行亚细胞定位预测。
1.3.5 高粱SBP基因启动子顺式作用元件分析
利用TBtools提取SBP基因的2000bp启动子序列,然后以fasta格式导入Plantcare(http://bioinformatics.psb.ugent.be/webtools/plantcare/ht-ml/)网站分析潜在的顺式作用元件,并统计各基序在.SBP启动子上的分布。
1.3.6 SBP基因家族共线性分析
利用TBtools软件分析高粱与拟南芥.SBP基因在染色体上的分布和共线性关系并作图,使用Adobe IllustratorCS5软件对图片进行优化。
1.3.7 总RNA的提取和实时荧光定量PCR(qRT-PCR)分析
用TIANCEN RNA Easy Fast植物组织RNA快速提取试剂盒提取总RNA,用TIAN-GEN FastKingCDNA第一链合成试剂盒反转录为CDNA,-20℃保存备用。用Premier 5.0设计qRT-PCR引物(表1),以SbACTIN为内参基因。PCR反应体系10μL:2xSYBRGreen5μL,上、下游引物各0.2μL,cDNA 0.5μL,ddH20 4.1μL。qRT-PCR程序:95℃30s;95℃Ss,58℃25s,72℃18s,循环40次:95℃Ss,650C1min。用2-△△Ct法进行基因表达量计算。
2 结果与分析
2.1 高粱SBP家族成员的鉴定
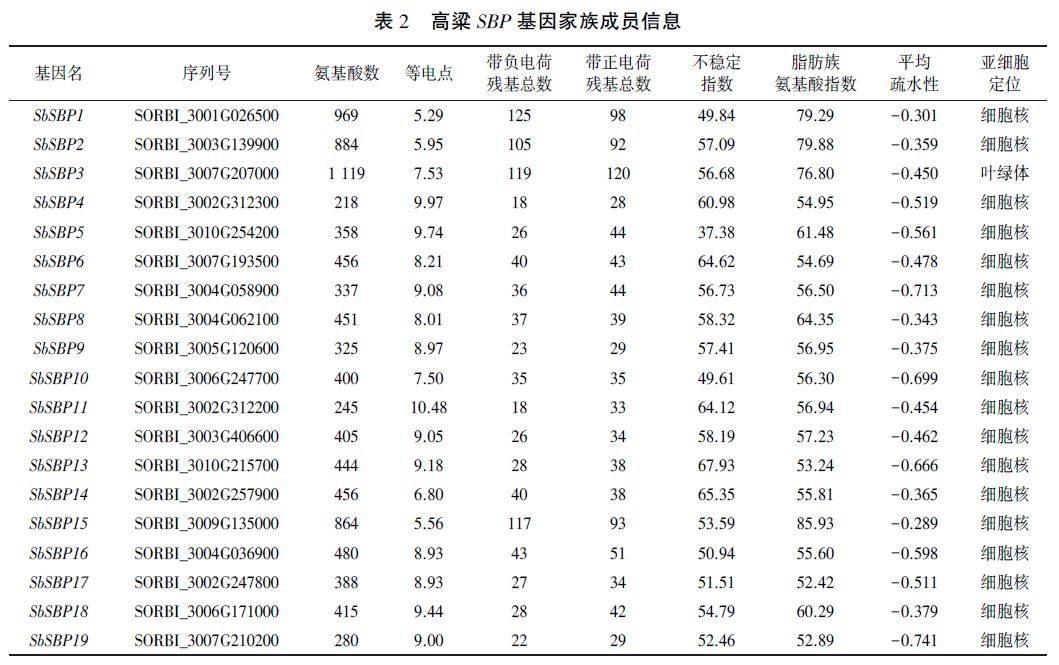
利用已发表的16个拟南芥SBP成员的氨基酸序列在Phytozome网站进行BLASTP序列比对,共获得21个高粱候选SBP基因。进一步通过Pfam和NCBI在线网站分析获得基因的蛋白结构域,去除不含SBP结构域的序列,最终得到19个高粱SBP家族成员(表2、图1)。
理化性质分析(表2)发现,高粱SBP家族成员的氨基酸序列长度差异较大,最短的仅218aa,最长的为SORBI_3007G207000,达1119aa。预测等电点差异也较为明显,其中酸性蛋白较少,仅有3条,碱性蛋白有16条,表明高粱SBP蛋白多为碱性蛋白。不稳定指数小于40的蛋白仅有SOR-BI_3010G254200,稳定性较高,其余蛋白的稳定性均较低。所有高粱SBP成员的平均疏水性均为负数,表明SBP蛋白都是亲水蛋白。亚细胞定位预测结果显示,除SORBI_3007G207000外,其余蛋白均定位在细胞核中,这可能与SBP转录因子行使功能有关。
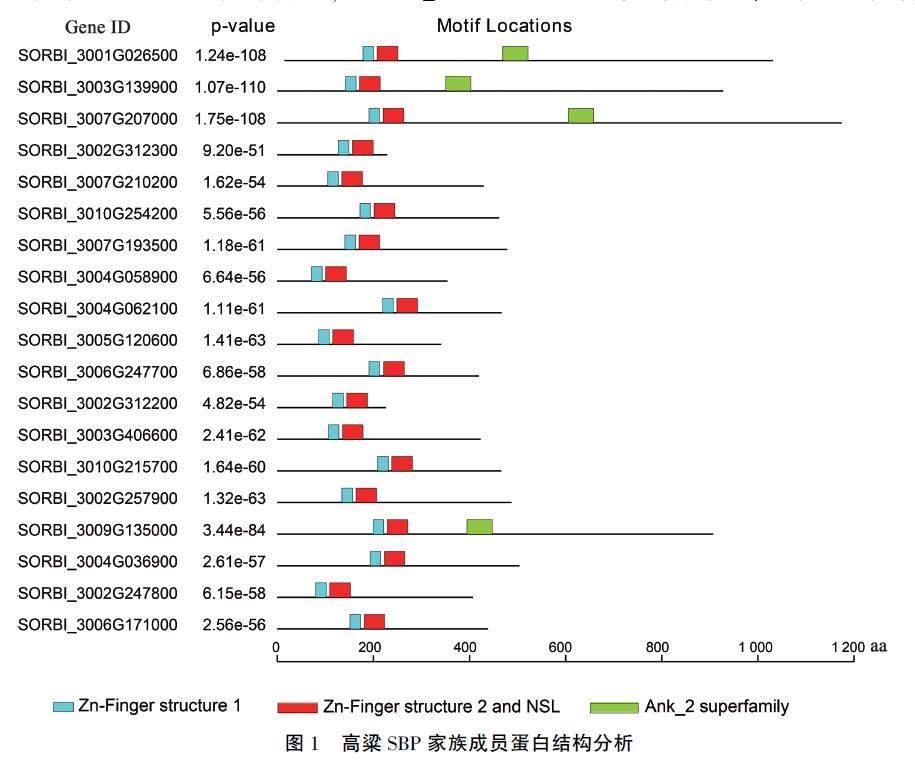
蛋白结构域分析结果(图1)表明,所有高粱SBP成员的氨基酸序列上均含有典型的SBP结构域,包括两个锌指结构和核定位信号。此外,SORBI_3001G026500、SORBI_3003G139900、SOR_BI_3007G207000及SORBI_3009G135000的氨基酸序列羧基端包含一个Ank_2 superfamily结构域,暗示着他们可能有一些共同的特殊功能。
2.2 高粱SBP家族系统发育分析
用拟南芥(16个)、水稻(19个)、玉米(28个)及高粱(19个)的SBP成员氨基酸序列,通过MEGA进行系统发育分析,结果将所有SBP大致分为3个亚类(图2),这与在水稻中的研究结果类似。每个亚类均包含水稻、拟南芥、玉米和高粱的SBP成员,表明SBP家族的分化在物种分化前即已形成。从进化枝上看,相对于拟南芥,高粱与水稻、玉米的SBP家族进化关系更为接近。
高粱的SBP成员也分为3类,分别含有7、5、7个SBP成员(图3)。DNA結构分析发现,在系统发育关系上比较靠近的基因,其外显子和内含子分布结构更为相似,如SORBI_3006G247700、SORBI_3004C062100和SORBI_3010C215700均包含3个外显子和2个内含子;SORBI_3001G026500、SORBI_3003G139900和SORBI_3007G207000分别含有10、10个和9个内含子。
2.3 高粱SBP家族成员的染色体定位分析
对高粱SBP基因的染色体定位分析发现,除第8条染色体外,其余9条染色体上均分布有SBP基因(图4)。其中,2号染色体上分布有4个SBP基因,数量最多,且表现出聚类分布的情况,两两聚在一起:4号和7号染色体上均分布有3个SBP基因,3、6、10号染色体上均有2个SBP基因,1、5、9号染色体上均仅有一个.SBP基因。
对高粱染色体的基因密度及其与拟南芥的共线性分析结果(图5)显示,高粱与拟南芥间存在61对共线性关系,并且高粱的SBP基因在染色体上主要位于基因高密度区域,表明SBP基因在高粱染色体上呈热点区域分布。
2.4 高粱SBP家族基因的启动子基序分析
为了解高粱SBP基因可能参与的生物学过程,对19个成员转录起始位点上游2000bp的启动子区域进行顺式作用元件预测,结果(表3)显示,高粱SBP基因启动子区拥有丰富的顺式作用元件,主要包含有脱落酸响应元件、厌氧诱导响应元件、茉莉酸甲酯响应元件、光响应元件、低温响应元件、MYB转录因子结合位点及昼夜节律调控位点等。这些顺式作用元件的存在暗示着SBP基因广泛参与了高粱的生物学过程。如除SORBI_3004G036900和SORBI_3002C312200外,其余17个成员启动子上均包含有脱落酸响应的ABRE结合基序,暗示着高粱SBP家族可能普遍参与了其干旱响应过程:茉莉酸甲酯响应元件TGACG在SORBI_300IG026500等15个成员启动子上均有分布,表明这些基因可能受到茉莉酸甲酯的诱导:此外,光响应元件G-box、MyB转录因子结合位点等在高粱SBP基因启动子上的分布也较为普遍。推测高粱SBP家族可能参与了植物响应干旱、光和低温等多种生物学过程。
2.5 高粱SBP家族基因的表达模式分析
为进一步研究高粱SBP基因的潜在功能,對19个SBP基因在高粱幼苗全株、叶、根、茎和花/种子中的表达模式进行了分析。通过Phytozome网站获得各个成员的组织表达量,并利用TBtools软件构建组织表达热图。结果(图6A)发现,各基因在高粱幼苗不同组织中的表达量存在明显差异,其中SORBI_3003G139900、SORBI_3007G207000、SORBI_300IG026500和SORBI_3009G135000在各组织中的表达量均最高,而SORBI_3006C171000、SORBI_3004G062100和SORBI_3010G215700在各组织中的表达丰度均较低。其中,SORBI_3003G139900、SORBI_3007G207000、SORBI_300IG026500等在茎中的表达量最高,SORBI_3007G193500在花/种子中的表达量最高,SORBI_3002G312200和SORBI_3002G312300在根中的表达量最低。表明这些SBP基因在高粱生长发育过程中发挥的功能具有差异性。
此外,本研究选取启动子区含有脱落酸响应元件ABRE的17个SBP基因检测其在干旱胁迫下的表达模式,结果(图6B)发现,这17个基因在干旱处理一段时间后,均呈现出表达量上调的趋势,暗示SBP家族基因参与高梁响应下旱胁迫的积极作用。其中SORBI_300IG026500、SORBI_3003G139900、SORBI_3004G058900、SORBI_3004_062100、SORBI_3005G120600、SORBI_3006G24770和SORBI_3009G135000均在干旱处理24h表达量最高,而SORBI_3010G254200、SORBI_3010G215700、SORBI_3002G247800、SOR-BI_3006G171000和SORBI_3007G210200在干旱处理36h表达量才达到最高。此外,SORBI_3010G254200和SORBI_3007G193500在干旱诱导初期先表现出抑制下调,随后才逐渐上调表达。可见,高粱的SBP家族成员随干旱胁迫时问的延长呈现出差异化的表达模式,暗示着其在响应干旱胁迫时可能有着不同的功能。
3 讨论与结论
作为植物特有的转录因子家族,当前的研究认为SBP家族参与了植物体内多种生物学过程,不仅参与了植物的生长发育调控,而且在植物响应逆境胁迫过程中发挥着重要作用。因此,开发植物SBP家族基因并进行功能研究具有重要意义。
不同物种的SBP基因数量存在显著差异,如矮牵牛(Petunia axillari.s)中有21个SBP基因,梅花(Prunus mume)中有15个SBP基因,白梨(Pyrus bretschneideri)中有32个.SBP基因。本研究从高粱中鉴定出l9个.SBP家族基因,与水稻的SBP基因数量相当,多于拟南芥,少于玉米。SBP基因数量的变化可能与物种的进化历史及环境选择相关。
本研究结果表明,高粱SBP基因的氨基酸序列均包含有典型且高度保守的SBP结构域,与在其他物种中的结果一致:部分基因的氨基酸序列羧基端包含一个Ank_2 superfamily结构域,该结构域为锚蛋白重复序列,但其在SBP行使功能过程中发挥的作用还未知。
系统发育分析发现,高粱、拟南芥、水稻、玉米的SBP家族成员可分为3个亚枝,并且每个亚枝中均含有这4个物种的SBP成员,表明SBP家族在单子叶和双子叶植物分化前即已扩张为3个亚枝,这与之前的研究结果一致。此外,高粱与水稻、玉米的SBP基因具有更近的进化关系,且同一进化枝的SBP直系同源基因可能具有类似功能。从系统发育树还可看出,拟南芥的SBP家族拥有更多的旁系同源基因对,如AT1 G20980.J与AT2CA7070.1,AT1G53160.1与AT3C15270.1等,表明在单双子叶物种分化后,高粱与拟南芥^SBP基因经历了不同的物种特异性基因复制或丢失事件。
有报道表明,SBP基因在植物响应非生物胁迫如干旱、盐胁迫等过程中发挥重要功能。如白桦(Betula platyphylla Suk.)SBP家族成员SPL9会受到盐和PEG-6000的诱导表达,拟南芥异源转基因实验发现SP/9通过清除活性氧提高植物对盐旱的耐受性:中国野生葡萄(Vitis)的SBP16在拟南芥中表达后通过调控SOS和ROS信号通路提高植株的干旱和盐抗性:水稻miR156k通过降低靶基因SPL3、SPL14和SPL17的表达减弱其抗冷性:玉米的部分SBP家族基因会受到干旱、冷、盐等的诱导表达。本研究发现,高粱19个SBP成员中有17个含有ABRE脱落酸响应元件,进一步的qRT-PCR检测显示,这17个基因受到干旱胁迫的诱导表达,但随胁迫时间延长的表达模式存在差异,结合这些基因在高粱不同组织中的差异化表达,推测它们可能在高梁响应干旱过程中发挥着不同的功能,这为后续探究SBP基因在高粱生长发育及非生物胁迫响应方面的作用机制奠定了一定的理论基础。
基金项目:贵州省科技计划项目(黔科合基础-ZK[2023]一般169);贵州省基层农技推广项目(仁怀基地示范[2301]01号);贵 州省科技成果应用及产业计划项目(黔科中引地[2022]4045)
猜你喜欢
中国中药杂志(2017年1期)2017-03-06
江苏农业科学(2016年8期)2017-02-15
安徽农学通报(2016年24期)2017-01-12
中国实用医药(2016年30期)2016-12-28
山东农业科学(2016年11期)2016-12-17
中国民族民间医药·上半月(2016年10期)2016-11-19
江苏农业科学(2016年4期)2016-06-14
现代园艺(2016年2期)2016-03-15
江苏农业科学(2015年11期)2016-01-27
江苏农业科学(2015年1期)2015-04-17
