
两个家蚕CPFL表皮蛋白基因的鉴定与表达分析
2024-06-11 00:55刘惠芬华丽峰刘文光娄齐年衣葵花李云芝郭光
山东农业科学 2024年3期
刘惠芬 华丽峰 刘文光 娄齐年 衣葵花 李云芝 郭光


摘要:昆虫CPFL表皮蛋白是一类重要的结构蛋白。本研究通过生物信息学分析从家蚕基因组中鉴定出2个CPFL基因,命名为BmHCP1和BmHCP2。氨基酸序列分析发现,2个蛋白在羧基末端均具有CPFL表皮蛋白典型的保守结构序列YCW以及重复序列AAH、AAPA、AAPV。进化树分析显示,2个蛋白与鳞翅目昆虫艺神袖蝶(Heliconius erato)和柑橘凤蝶(Papilio xuthus)具有较高同源性。荧光定量PCR分析显示,BmHCP1和BmHCP2存在组织、时相特异性表达,幼虫期在头部、中肠、表皮等组织中表达量较高;蛹期除翅原基、血淋巴、触角和足等组织外均有较高表达量:成虫期在足、触角、翅原基、表皮等组织中表达量较高;蛹期在大多数组织的表达量明显高于幼虫期和成虫期。研究结果为进一步探明家蚕表皮蛋白基因的功能奠定了基础。
关键词:家蚕;CPFL表皮蛋白;序列分析;表达分析;荧光定量PCR
中图分类号:S881.2:Q781 文献标识号:A 文章编号:1001-4942(2024)03-0139-06
表皮对昆虫适应复杂多变的生存环境至关重要,具有皮肤和外骨骼的双重作用,主要成分是几丁质和表皮蛋白。昆虫表皮蛋白属于结构蛋白,按照保守结构域可分为12个家族,包括R&R、Tweedle、CPF和CPFL等。R&R表皮蛋白家族最早由Rebers等发现,含有GCYAEGYPP保守结构域,已公布的521种节肢动物的表皮蛋白中,2/3以上具有该保守结构域,并已证实其具有结合几丁质的作用。Tweedle表皮蛋白家族是研究果蝇(Drosophila melanogaster)表皮表型及形态发生机制时偶然发现的,该蛋白家族基因发生突变会导致果蝇表皮出现各种突变性状。CPF是一类小的表皮蛋白家族,具有51个保守氨基酸的表皮蛋白基序,其保守结构域是从黄粉虫(Tenebrio mlitor)和东亚飞蝗(Locustamigratoria)的6个表皮蛋白序列中被识别JLH来的,另外,CPF表皮蛋白家族还具有保守的羧基端YGW、YAW、HGW序列。CPFL是一种具有类似于CPF保守基序的表皮蛋白家族,两者相似性很高,有许多共同的保守基序,尤其是羧基端保守序列。昆虫表皮蛋白基因的表达受环境、激素、转录因子和内含子等因素共同影响,具有时相和组织特异性。昆虫表皮蛋白及其编码基因被认为是研究昆虫蜕皮和变态的调控机理、理解昆虫发育期角质层在生理生化及结构上的修饰的重要靶标。
Futahashi等研究发现,家蚕存在220个表皮蛋白基因,其中包括148个R&R、4个Tweeclle、1个CPF和4个CPFL表皮蛋白基因。梁九波在家蚕基因组中鉴定到255个表皮蛋白基因,同样包含4个CPFL表皮蛋白基因。Futahashi等发现凤蝶幼虫中存在2个硬表皮蛋白基因,HCP1和HCP2(GenBank登录号分别为AB264673、AB264675),与凤蝶拟态特殊的突起结构密切相关。本研究利用上述2个凤蝶硬表皮蛋白基因序列,通过生物信息学分析,得到2个家蚕表皮蛋白基因(BmHCP1,BmHCP2),并在NCBI注册,GenBank登录号分别为FJ577507和FJ577508。同时,对这2个基因进行时相和组织表达分析,以期为探明家蚕表皮蛋白基因的功能奠定理论基础。
1 材料与方法
1.1 试验材料
供试家蚕品种为JY-I,由中国农业科学院蚕业研究所提供,常规饲养。
1.2 主要试剂
各种限制性内切酶,T4 DNA连接酶购自Pro-mega公司;pMD18-T载体购自TaKaRa公司;DL8000 Plus DNA Marker购自北京全式金生物技术有限公司:引物合成和DNA测序委托北京三博远志生物技术有限责任公司完成。
1.3 生物信息学分析
利用NCBI网站(https://www.ncbi.nlm.nihgov/)公布的2条柑橘风蝶硬表皮蛋白基因的序列(GenBank登录号:AB264673,AB264675),进行家蚕tBIASTx序列比对,得到EST序列,利用DNAStar软件进行装配并手工校正,获得家蚕表皮蛋白基因序列,利用SignaIP 5.0(https://serv-ices.healthtech.dtu.dk/service.php?SignaIP-5.0)在线工具对其信号肽进行分析,利用ClustaIX1.83软件(默认设置)进行多序列比对,利用MEGA 6.0软件中的邻接法构建系统进化树。
1.4 反转录PCR
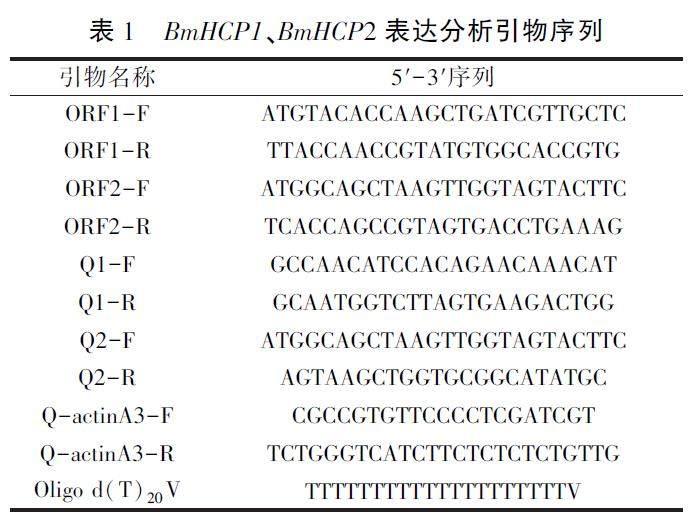
利用TRIZol试剂盒提取家蚕不同发育时期、不同组织的总RNA,紫外分光光度法测定总RNA浓度。以Oligo(dT)20V为引物进行反转录得到cDNA;反应条件为30℃10min,42℃1h,70℃15min,4℃5min。根据1.3拼接得到的cDNA序列,设计特异性引物(表1),以cDNA为模板(约相当于10ng总RNA转录量)进行反转录PCR。PCR反应程序:95℃预变性5min:55℃退火40s,72℃延伸1min,30个循环:72℃补延伸10min。PCR产物经琼脂糖凝胶电泳和测序检测,详细方法参照文献[20-21],每個样品重复3次。
1.5 Real-time PCR和标准曲线制作
参照1.4提取家蚕不同时期、不同组织的总RNA,取1μg总RNA进行反转录,以所得cDNA为模板(约相当于10ng总RNA转录量),以家蚕actinA3基因为内参基因,进行Real-time PCR,每个样本至少3次重复。反应条件为:95℃3min;95℃20s,58℃20s,72℃30s,40个循环:熔解曲线从55℃到95℃,每0.5℃读取10s。
标准曲线的制作:将包含日的基因及内参基因片段克隆到pMD18-T载体上,测定质粒浓度并按10s-104拷贝μL进行梯度稀释作为标准样进行Real-time PCR。以所得Ct值对相应拷贝数的对数值作图,即得各个基因扩增的标准曲线及其线性方程v=ax+b。对Real-time PCR结果进行绝对定量分析,根据标准曲线方程计算x值,则待测基因的拷贝数Y=10x。
2 结果与分析
2.1 BmHCP1、BmHCP2序列号分析
利用2条柑橘凤蝶硬表皮蛋白基因序列在NCBI中进行家蚕tBIASTx序列比对,获得2个家蚕表皮蛋白基因BmHCPl(CenBank:FJ577507)和BmHCP2(GenBank:FJ577508),均位于第23号染色体上,位置为NC_051380.1(10459128-10460263)和NC_051380.1(10437133-10438764),编码区(CDS)序列长度分别为918bp和714bp,分别编码305个和237个氨基酸。对其氨基酸序列进行分析,结果(图1)表明,2个基因编码的蛋白在羧基末端均具有CPFL表皮蛋白典型的保守氨基酸序列YCW,且在其内部有许多重复的短的片段:AAH、AAPA、AAPV等。用signaIP 5.0在线预测软件对2个基因的氨基酸序列进行信号肽及其酶切位点分析,结果显示,2个表皮蛋白均具有信号肽,酶切位点均位于第17个和18个氨基酸之间,分别是VRA-GG和ARA-GG。
2.2 几种昆虫CPFL蛋白氨基酸序列比对分析
将BmHCP1和BmHCP2蛋白羧基端序列与鞘翅日昆虫蔗根象鼻虫(Diaprepes abbreviatus),鳞翅目昆虫柑橘凤蝶(Papilio xuthus)、艺神袖蝶(Heli-conius erato),双翅目昆虫冈比亚按蚊(Anopheles gambiae)、埃及伊蚊(Aedes aegypti),直翅目昆虫东亚飞蝗(Locusta migratoria)的CPFL表皮蛋白的羧基端序列(登录号见表2)利用ClustaIX l.83软件进行多序列比对,发现上述几个目昆虫的CPFL蛋白羧基端具有较高保守性,除艺神袖蝶HeCPFL2外均含有YGW/YAW/HGW序列(图2)。
2.3 CPFL蛋白家族系统发生分析
利用MEGA 6.0软件将BmHCPI和BmHCP2蛋白序列与已知的鞘翅目、鳞翅目、双翅目、直翅目昆虫的CPFL表皮蛋白序列进行比对,采用邻接法(NJ)构建系统进化树。结果显示,2个家蚕表皮蛋白与鳞翅目昆虫艺神袖蝶和柑橘凤蝶的CPFL蛋白家族位于一个大的进化分枝上,2个家蚕表皮蛋白并不在同一个亚进化分支上(图3)。
2.4 BmHCP1、BmHCP2的定量表达分析
对家蚕幼虫(5龄第3天)、蛹(第3天)、成虫(第2天)三个时期的头部、表皮、脂肪体、丝腺、马氏管、翅原基、精巢、卵巢、中肠、气管丛、血淋巴、触角等组织中BmHCP1和BmHCP2基因表达量进行分析。结果(图4、图5)显示,两基因表达均存在时相和组织特异性,蛹期在大多数组织中的表达量明显高于幼虫期和成虫期:幼虫期Bm-HCP1在头、中肠、表皮等组织中表达量相对较高,BmHCP2在头、精巢、中肠等组织中表达量相对较高:蛹期除在翅原基、血淋巴、触角和足中表达量较低外,在其他组织中表达量相对较高:成虫期在翅原基、表皮、足、触角等组织中表达量较高。幼虫期多数组织中,BmHCP2的表达量明显高于BmHCP1;蛹期多数组织中BmHCP1和BmHCP2表达量相当:成虫期触角和足中BmHCP2的表达量明显高于BmHCP1。
3 讨论与结论
昆虫的表皮由几丁质和表皮蛋白组成,随着家蚕、按蚊、果蝇、蜜蜂和赤拟谷盗等昆虫全基因组测序相继完成,对表皮蛋白的研究也随之展开。表皮蛋白是研究昆虫蜕皮与变态调控机制的重要模型,受到越来越多的重视。本研究利用柑橘凤蝶幼虫硬表皮蛋白基因序列,通过生物信息学分析获得了2个家蚕表皮蛋白基因BmHCP1和Bm-HCP2,经序列分析发现均属于CPFL蛋白家族。Futahashi等在家蚕表皮蛋白基因组研究中鉴定了4个编码CPFL蛋白的基因,它们形成排列紧密的表皮蛋白串联基因簇,其中BomrCPFL3(BR000420)和BomrCPFL4(BR000421)与本研究中发现的BmHCP1和BmHCP2仅有少数几个氨基酸的差异,且BmHCP1和BmHCP2也串聯分布在同一条染色体上,进一步确定BmHCP1和Bm-HCP2属于CPFL蛋白家族基因。在昆虫领域,家蚕、果蝇、按蚊等都具有众多基因家族编码表皮蛋白,其中对按蚊表皮蛋白基因研究的较为通透,它们同样以排列紧密的串联基因簇形式存在,其氨基酸序列几乎完全一样。
本研究对家蚕BmHCP1和BmHCP2基因进行时相、组织表达分析,结果显示,2个基因在家蚕不同发育时期、不同组织中存在特异性表达,且在蛹期的表达量明显高于幼虫和成虫期。Fu-tahashi等在家蚕吐丝后发现了CPFL转录因子,推测其可能与家蚕蛹期硬表皮刚性结构的形成有关:还发现,即使是相同基因簇的相邻表皮蛋白基因,也存在组织和时相特异性表达,这与本研究结果相似。
家蚕表皮蛋白基因BmCPG2、BmCPR2、Bm-CPR63等与鳞毛的构建有关。赤拟谷盗表皮蛋白基因TcCPAP3-B缺失后,足的股骨-胫骨关节以及后足的股骨-胫骨关节和胫骨-跗骨关节变得刚硬并畸形,推测该基因可能参与足的构建。2个凤蝶硬表皮蛋白基因与其拟态特殊的突起结构密切相关。本研究荧光定量分析发现,BmHCP1和BmHCP2在家蚕成虫期翅、触角及足中表达量较高,推测可能参与家蚕翅、触角及足刚性结构的形成,具体功能有待进一步研究。
BmHCP1和BmHCP2的生物信息学分析及其时相、组织表达分析为家蚕表皮蛋白基因的研究奠定了良好的基础,今后可进一步开展BmH-CP1和BmHCP2上游调控元件如核心启动子、起始子、基因表达调控元件以及调控模式等方面的研究。
基金项目:山东省蚕桑产业技术体系建设项目(SDAIT-18-02,SDAIT-18-03);山东省农业科学院农业科技创新工程项目(CXGC2023C03);国家现代农业产业技术体系烟台综合试验站项目(CARS-18-SY208)
猜你喜欢
江苏农业科学(2016年11期)2017-03-21
江苏农业科学(2017年1期)2017-02-27
山东农业科学(2016年11期)2016-12-17
中国中药杂志(2016年20期)2016-11-19
现代养生·下半月(2015年8期)2015-11-16
江苏农业科学(2015年5期)2015-10-20
江苏农业科学(2015年9期)2015-10-20
江苏农业科学(2015年9期)2015-10-20
江苏农业科学(2015年8期)2015-09-10
江苏农业科学(2015年8期)2015-09-10
