
低温环境下酪氨酸和半胱氨酸联合刺芒柄花素调控小鼠肝脏产热的机制
2024-04-01 05:27李仕韦徐彬张博熙李士泽
黑龙江八一农垦大学学报 2024年1期
李仕韦,徐彬,张博熙,李士泽
(黑龙江八一农垦大学动物科技学院,大庆 163319)
中国北方地区纬度较高、白昼短,临近冬季风发源地,冬季寒冷且漫长,常导致畜禽暴露于低温环境中。冷暴露与机体健康密切相关,适当的低温环境对于某些种植业的发展具有一定优势,但对于养殖业来说恰恰相反。在寒冷地区的冬季、春初或秋末,由于气温变化幅度大,畜禽出现冷应激现象的几率非常高。研究上,将机体对寒冷刺激做出的非特异性应答反应,称之为冷应激。冷应激可对畜禽的健康和生产力造成不利影响,导致家畜生长缓慢、生产效能下降、饲养效率降低和免疫功能障碍[1]。当畜禽处在冷应激状态下时,为保持机体正常的生存体征,机体中的能量首先被用于保持体温,这就导致没有多余的能量来促进机体生长发育,进而导致动物体重下降、增长缓慢甚至会引发多种疾病。综上可知,冷应激已成为制约我国北方畜牧业发展的重要因素之一。
机体在感受环境中的冷热温度刺激后,自身可以进行体温调节,从而将体温控制在一定范围内以适应环境的变化。其中产热过程主要通过肝脏、棕色脂肪组织(BAT)、骨骼肌等组织及器官在生理代谢及运动做功过程中释放能量来实现,可分为非战栗产热(non-shivering thermogenesis,NST)和战栗产热(shivering thermogenesis)。质子漏产热作为NST 的三大途径之一,主要发生在BAT、肝脏、骨骼肌和肾等组织器官中。线粒体可以通过三羧酸循环氧化机体的营养物质,电子传递链泵出的H+在线粒体内膜上所形成的浓度差和电位差用于储存氧化所得的能量。当线粒体内膜上存在质子漏时,H+会通过质子漏泄漏回线粒体基质中,此过程降低电位差并且不伴随ATP 合成,因此将电势能转化为了热能。生热过程中的一个关键角色是解偶联蛋白-1(UCP1),它将ATP 合成与氧化磷酸化解偶联,将后者过程中产生的质子直接转回线粒体内膜,进而产生热量[2-3]。啮齿动物中的BAT 比人类更丰富,它在冷暴露中的激活有助于全身脂质清除和血糖控制,但在冷暴露后啮齿动物产热调节的研究方面大量学者很少考虑到BAT 以外的器官。肝脏作为NST 的主要产热组织器官之一,也是机体暴露在冷环境下的重要产热组织,但其生理功能有局限性,故在冷环境中NST 产热增加量也受到限制,因此促进冷环境下肝脏组织产热能力对动物机体维持体温恒定有着重要的意义。
酪氨酸是一种非必需氨基酸,也是一种抗氧化剂,能清除体内的自由基并抑制脂质过氧化。半胱氨酸作为谷胱甘肽的前体物质,具有创伤修复,抗菌消炎和解毒的功能[4-6]。刺芒柄花素[7-羟基-3(4-甲氧基苯基)色酮]是一种异黄酮化合物,广泛存在于天然植物中,它是多种中草药中的主要生物活性化合物之一。已有研究表明,刺芒柄花素具有抗氧化[7]、抗炎[8]、抗细胞凋亡[9]等作用。研究发现,刺芒柄花素通过TFEB 介导的溶酶体生物合成改善肝脏脂肪变性[10];黄芪组分刺芒柄花素作为核激素受体PPARγ 的激动剂,能够有效调控白色脂肪细胞褐色化,减少肥胖及其相关代谢疾病的发生[11]。新合成的刺芒柄花素类似物XH601 可降低高脂血症小鼠的脂肪重量和血清甘油三酯、总胆固醇和低密度脂蛋白水平,同时增加血清高密度脂蛋白水平[12]。
刺芒柄花素在临床中具有稳定性好、副作用小等特点。介于上述背景,刺芒柄花素在冷暴露中尚未有研究。因此,试验选择刺芒柄花素为治疗药物,探究其在冷暴露条件下联合酪氨酸和半胱氨酸对小鼠肝脏产热能力的影响,为提高畜禽冬季抗冷应激能力提供理论依据。
1 材料与方法
1.1 主要试剂及仪器
主要试剂:4%多聚甲醛、苏木精染色剂、伊红染色剂、油红O 染色试剂盒、RIPA 裂解液、PMSF 蛋白酶抑制剂、BCA 蛋白浓度测定试剂盒、SDS-PAGE 制胶试剂盒、SDA-PAGE 蛋白上样缓冲液(5X)均购自上海碧云天生物技术有限公司;蛋白marker、底物发光液、PVDF 膜购自美国MILLIPORE 公司;PGC-1α鼠单克隆抗体、PPAR-γ 兔多克隆抗体、UCP1 兔多克隆抗体、β-Tubulin 鼠单克隆抗体、HRP 标记的山羊抗兔IgG 和山羊抗鼠IgG 均购自美国proteintech 公司。
主要仪器:37 ℃恒温培养箱,台式低速离心机,低温离心机,倒置生物显微镜,制冰机,-80 ℃超低温冰箱,4 ℃冰箱,-20 ℃冰箱,MM400 混合型球磨仪,酶标仪,电泳槽,转膜仪,摇床,电子天平。
1.2 实验动物
4 周龄雄性SPF 级C57BL/6J 小鼠购自辽宁长生生物技术股份有限公司。将其随机分为常温对照组(RT 组)、冷暴露组(CE 组)、冷暴露+8 mg/kg/d 刺芒柄花素组(CE+FMN8 组)及冷暴露+16 mg/kg/d 刺芒柄花素组(CE+FMN16 组),每组10 只,昼夜交替光照12 h,自由饮水,室温(26±1)℃预饲一周后对加药组小鼠进行相应浓度的灌胃,灌胃结束后与冷暴露组一起每天置于4 ℃人工气候室中冷刺激3 h,为期28 d。4 周后处死小鼠并采集肝脏组织。
1.3 实验饲料
半胱氨酸纯度99.66%,酪氨酸纯度>98%,购自浙江一诺生物科技有限公司。在1 kg 鼠粮中添加60 g的酪氨酸混合半胱氨酸(1∶1 混合),加工自沈阳前民饲料厂。
1.4 H&E 染色
将肝脏组织固定于4%多聚甲醛缓冲液,置于包埋盒中,流水冲洗30 min,利用不同浓度的酒精脱水,在二甲苯溶液中使组织透明后,将透明组织放在石蜡中,包埋后进行切片。染色前,采用二甲苯溶液脱蜡后,放入由高到低浓度梯度的酒精中,用蒸馏水清洗。将切片置于苏木精染液中几分钟,放于盐酸乙醇和氨水中各30 s 进行分色,用伊红染色液染色2~3 min,通过不同浓度的酒精脱水,利用二甲苯溶液透明后封片。在200 放大倍率下观察各组肝脏组织病理形态。
1.5 免疫组织化学技术
将固定后的肝脏组织进行固定和脱水处理,将处理后的组织包埋在石蜡中,包埋后的肝脏组织用切片机切成5 μm 厚的切片,将处理后的切片进行脱蜡和抗原修复。处理完毕后,对切片进行血清封闭然后加一抗4 ℃孵育过夜。一抗孵育结束后加入二抗37 ℃孵育30 min,二抗孵育结束后加入SABC 37 ℃孵育30 min,孵育结束后加入显色剂,将显色后的切片加入苏木精复染,复染结束后进行脱水和封片。
1.6 产热因子表达水平的检测
用RIPA 提取肝脏组织中的总蛋白质,提取后在4 ℃和12 000 RCF 的条件下离心10 min。离心结束后收集上清液用BCA 蛋白浓度测定试剂盒测定蛋白浓度,加入上样缓冲液使其变性。以上试剂均购自碧云天生物技术有限公司。将每个样品等量的加入SDS-PAGE 凝胶上并进行电泳。电泳后,将蛋白转移至PVDF 膜上,用5%脱脂奶粉封闭PVDF 膜2 h,封闭结束后孵育一抗(1∶1 000)8 小时,加入HRP标记的山羊抗兔和山羊抗鼠(1∶10 000 稀释)孵育1 h。用Chemi Doc XRS+(Bio-Rad,USA)凝胶成像系统检查印迹,并用Image-J 软件分析目的蛋白与内参的灰度比值。
1.7 统计学分析
所有数据使用统计软件SPSS 26.0 进行分析,数据用平均值±标准误差(Mean±SEM)表示,组间Western Blot 结果比较分析用 One-way ANOVA,P<0.05 具有统计学意义。
2 结果
2.1 小鼠肝脏组织病理变化
肝脏组织形态是评价肝损伤的重要方法和依据,图1 为小鼠肝脏HE 染色结果图。通过HE 染色可以发现,RT 组小鼠肝脏组织结构相近,细胞核大小相似,分布均匀,肝细胞轮廓清晰,肝索自中央静脉呈放射状排列。CE 组肝组织出现炎性细胞浸润,肝细胞出现点状坏死,并且有部分细胞出现核皱缩。CE+FMN16 组肝索结构清晰,未见明显炎性细胞浸润和肝细胞坏死。由此可以判断慢性冷暴露诱导了小鼠的肝损伤,这种损伤可以被酪氨酸和半胱氨酸联合刺芒柄花素所缓解。
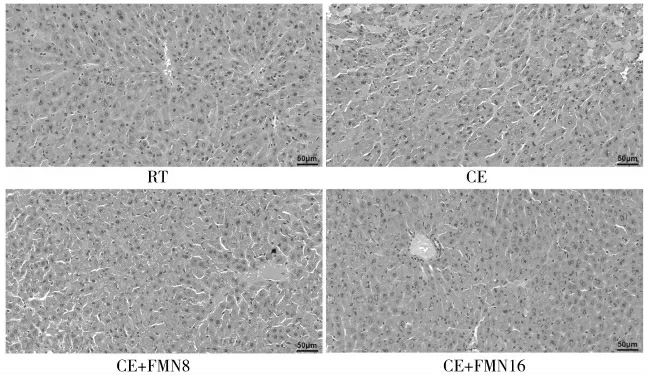
图1 小鼠肝脏组织HE 染色形态图(200×)Fig.1 HE staining morphology of liver tissue in mice(200×)
2.2 UCP1 的IHC 结果
为探究慢性冷暴露对小鼠肝脏产热能力的影响,研究采用免疫组化法对肝组织UCP1 蛋白的表达量进行量化统计。免疫组化结果显示,相比于RT 组,CE 组中肝脏UCP1 阳性染色表达量显著降低(图2);与CE 组相比,CE+FMN16 组中肝脏UCP1 阳性染色表达量显著升高,表明冷暴露条件下酪氨酸和半胱氨酸联合刺芒柄花素后,小鼠肝脏组织中的UCP1 表达量显著增强,进而促进小鼠肝脏组织发挥产热作用。

图2 小鼠肝脏组织IHC 结果图(200×)Fig.2 IHC results of mouse liver tissue(200×)
2.3 产热因子表达水平
通过Western Blot 方法检测产热相关因子的表达水平,结果如图3 所示。与RT 组相比,慢性冷暴露对小鼠肝组织中PGC-1α 和UCP1 的表达水平无显著影响,但显著上调了PPAR-γ 的表达;与CE 组相比,酪氨酸和半胱氨酸联合16 mg/kg/d 刺芒柄花素显著上调了产热因子PGC-1α、PPAR-γ 和UCP1 的表达水平。综上所述,添加刺芒柄花素可以显著增强肝脏产热相关蛋白的表达,从而促进肝脏产热来维持体温。

图3 Western blot 检测各组小鼠肝脏中产热蛋白的表达水平Fig.3 Western blot detection of the expression levels of thermogenic proteins in the liver of each group of mice
3 讨论
寒冷是北方地区动物面临的最常见的应激因素之一,寒冷因素给机体带来的损伤是不可估量的。动物长期处于寒冷环境中,会破坏机体内环境的平衡状态,同时也会增加机体的产热,降低动物的生产性能和饲料利用效率,严重制约了畜牧业的发展。在寒冷环境下,细胞、器官会协调一致的维持体温恒定,以平衡内环境稳态[13-14],肝脏是机体暴露在冷环境下的重要产热组织,但其生理功能有局限性,故在冷环境中NST 的产热增加量也受到限制,因此促进冷环境下肝脏组织产热对动物机体抗冷应激进而维持体温恒定有着重要的意义。
寒冷引起的压力和细胞反应,通常被称为适应性产热,对于维持体温从而确保正常的器官功能至关重要。产热程序的表达是由一个复杂的转录机制调控的,它涉及到大量的转录因子、共同激活物和共同抑制物。最早发现适应性产热的转录调控因子之一是过氧化物酶体增殖物激活受体γ(PPARγ)共激活因子1-α(PGC1α)。PGC1α 在冷暴露、运动或禁食时被诱导,主要通过核受体PPARγ 发挥适应性产热作用,也可通过甲状腺激素受体发挥作用[15-17]。最近研究表明,PGC-1α 在皮下白色脂肪组织中对β-肾上腺素信号和冷暴露等生热刺激有诱导作用,并通过与关键转录因子PPAR-γ 结合刺激生热基因的表达,促进白色脂肪细胞向米色脂肪细胞的转化[18]。此外,在白色脂肪细胞中过表达PGC1α 可诱导UCP1和呼吸链关键线粒体酶的表达[15]。相反,小鼠脂肪特异性PGC1α 缺失导致冷暴露后的产热反应减弱,核心体温降低,脂肪库中产热基因表达减弱。研究发现,酪氨酸和半胱氨酸联合刺芒柄花素的处理增加了小鼠肝脏中产热因子PGC-1α、PPAR-γ 和UCP1的表达,进而增强小鼠肝脏的产热能力。
UCP1 是冷介导的产热的关键调节因子,也是非颤栗性产热的标志蛋白[19-21]。研究发现,在冷敏感试验中,缺乏UCP1 基因的小鼠体温下降显著,而具有UCP1 基因的正常小鼠能维持机体正常体温,这说明冷刺激下小鼠体温主要通过UCP1 蛋白产热来维持[22]。PGC-1α 作为一个共激活因子,可以与PPARγ相互作用,PPARγ 可以结合到UCP1 启动子区域,调控UCP1 的表达。UCP1 可以将氧化呼吸电子传递链产生的质子动力势能转化为热量,增加非战栗产热消耗能量[23-24]。通常情况下,机体的体温调节依赖于中枢神经系统的调控和外周效应器的生热作用。在研究酪氨酸和半胱氨酸联合刺芒柄花素增强冷环境下小鼠肝脏产热能力的作用机理方面,试验从效应器着手,分析了肝脏产热组织的产热因子表达变化情况,研究发现,酪氨酸和半胱氨酸联合刺芒柄花素可以激活肝脏组织内UCP1 蛋白的表达,并最终促进机体生热。
4 结论
研究表明,日粮中补充氨基酸可以提高肝脏组织产热的能力,对机体起到保护作用。冷暴露下酪氨酸和半胱氨酸联合刺芒柄花素通过增强产热因子PGC-1α、PPARγ 和UCP1 的表达水平,促进小鼠肝脏产热,促进能量输出,为提高畜禽的抗冷应激能力提供一定的理论基础。
猜你喜欢
森林工程(2020年6期)2020-12-14
江苏农业科学(2020年8期)2020-06-01
中成药(2018年10期)2018-10-26
中成药(2018年9期)2018-10-09
天然产物研究与开发(2018年1期)2018-02-02
中成药(2018年1期)2018-02-02
山西中医药大学学报(2017年5期)2017-04-04
中国塑料(2016年7期)2016-04-16
云南师范大学学报(自然科学版)(2015年5期)2015-12-26
电源技术(2015年7期)2015-08-22
