
基于线粒体cox1、nad1 和rrnL 基因探讨马圆线科线虫间的亲缘关系
2024-04-01 05:27:24张中怀高远兰卓邱阳元邱鸿宇安琪高俊峰王春仁
黑龙江八一农垦大学学报 2024年1期
张中怀,高远,兰卓,邱阳元,邱鸿宇,安琪,高俊峰,王春仁
(1.黑龙江八一农垦大学动物科技学院,大庆 163319;2.台州学院生命科学技术学院)
马作为家畜的重要组成部分,是推动人类文明进步的重要载体,在人类社会发展史上发挥了其他家养动物无法比拟的作用[1]。马体内的寄生线虫不仅数量大,而且种类多,是影响马匹健康的重要因素,其中数量最大的一类线虫就是圆线科(Strongylidae)线虫,又称为马圆线虫。这类线虫呈世界性分布,在我国马群中也普遍存在[2-5]。当马圆线虫大量寄生时会造成宿主的贫血、消瘦,甚至死亡,给养马业带来严重的经济损失[6]。
马圆线虫主要包括盅口亚科(Cyathostominae)和圆线亚科(Strongylinae),共涉及19 个属和64 个种[7]。近年来,随着分子生物学的迅速发展,人们对于圆线科线虫的分子分类和亲缘关系有了更深入的了解,也改变了对传统分类的认识。线粒体DNA 具有结构简单、在细胞中大量存在、多为母系遗传、缺少重组和内含子以及较高的进化速率等特点,因此作为一种分子标记被广泛用于隐存种确定、分子进化、种群遗传、物种鉴定和不同分类水平上的系统发生关系研究[8-10]。Gao 等[9]基于线粒体全基因组数据证明,基于形态学而被归类为圆线亚科的三齿线虫与盅口亚科线虫亲缘关系更近。通过对来自不同地点微小杯冠线虫线粒体基因组数据分析发现,微小杯冠线虫是一个至少有三个不同种的复杂种[10]。
虽然马的圆线虫有64 种,但开展分子研究的并不多。因此,研究对新分离的黑龙江省大庆市马肠道的长伞杯冠线虫(Cylicostephanus longibursatum)、冠状冠环线虫(Coronocyclus coronatus)、细口杯环线虫(Cylicocyclus leptostomus)、拉氏杯口线虫(Poteriostomum ratzii)、麦氏副杯口线虫(Parapoteriostomum mettami)和无齿圆形线虫(Strongylus edentatus)6 种圆线虫的线粒体细胞色素c 氧化酶亚基Ⅰ(cox1)、烟酰胺腺嘌呤二核苷酸脱氢酶亚基Ⅰ(nad1)和核糖体大亚基(rrnL)进行扩增、测序和比较分析,然后以三个基因串联序列为标记基因,采用贝叶斯法(Bayesian inference,BI)法构建进化树来探讨圆线科线虫间的亲缘关系,为进一步研究圆线科线虫的分类及遗传进化关系提供基础资料。
1 材料和方法
1.1 虫体来源
在大庆某屠宰场收集马肠道粪便,反复水洗沉淀收集虫体,再通过生理盐水清洗3 次,置于显微镜下观察,参照文献进行形态学鉴定,将初步鉴定的长伞杯冠线虫、冠状冠环线虫、细口杯环线虫、拉氏杯口线虫、麦氏副杯口线虫和无齿圆形线虫保存于70%酒精中备用。为保证虫体鉴定的准确性,再以6种虫体的DNA 为模板,扩增其ITS 序列(引物见表1),通过序列分析鉴定到种。

表1 引物信息及PCR 扩增条件Table 1 Primer information and PCR amplification conditions
1.2 DNA 的提取及目的基因的扩增
1.2.1 DNA 的提取
虫体DNA 根据组织基因提取试剂盒(天根公司,北京)说明书进行提取。每种线虫取虫体1 条,在蒸馏水中反复冲洗3 次,加入200 μL GA 缓冲液和20 μL 蛋白酶K,置于56 ℃水浴锅中2~3 h,直到虫体完全溶解,期间要每隔30 min 摇晃一次。待虫体完全溶解后取出,加入200 μL GB 缓冲液,混匀后于70 ℃水浴锅中放置10 min,然后于4 ℃放置3 min,取出后加入-20 ℃的无水乙醇200 μL,充分颠倒混匀后瞬离。将上述所有的液体加入试剂盒内的吸附柱中,12 000 r·min-1离心30 s,弃废液。然后加入500 μL GD 缓冲液,12 000 r·min-1离心30 s,弃废液;再加入700 μL PW 缓冲液,12 000 r·min-1离心30 s,弃废液,此步骤需重复一次。加完缓冲液后,12 000 r·min-1离心2 min,然后置于室温3 min,晾干残余的液体。将吸附柱置于1.5 mL 离心管中,加入100 μL 的洗脱液TE,室温放置3 min 后12 000 r·min-1离心2 min,-20 ℃保存备用。
1.2.2 目的基因的克隆及测序
参考先前文献报道[6,11]设计引物扩增6 种线虫cox1、nad1 和rnnL 基因,引物序列由北京擎科生物工程技术有限公司合成。PCR 反应的总体系为25 μL,具体如下:1 μL DNA 模板,2.5 μL ExTaq buffer(pH 8.5),2 μL dNTP mixture(2.5 mM),0.5 μL 引 物(10 pmol·mL-1)和0.2 μL ExTaq DNA 酶(5 U·mL-1)。扩增程序如下:94 ℃预变性5 min;94 ℃变性1 min,55 ℃退火1 min,72 ℃延伸1 min,35 个循环;72 ℃延伸7 min(表1)。扩增产物经琼脂糖凝胶电泳检测,凝胶成像系统成像。取含有目的片段的凝胶置于1.5 mL 离心管中,并标注相应的编号,按照胶回收试剂盒(天根)说明书的步骤进行胶回收,纯化后的DNA 产物与PMD-18T 载体在16 ℃连接12 h 后用于克隆,将克隆阳性菌液委托吉林库美测序公司进行测序。
1.3 序列分析
测得的目的序列利用DNAstar 5.0 软件中的Editseq 程序进行序列编辑,从GenBank 数据库中获取其他圆线科线虫相应基因序列(表2)。对A+T 含量和AT 斜率进行计算,用ClustalX 1.83 软件和DNAstar 5.0 中的Align 程序进行DNA 同源序列排列,并人工核对校正。所得序列与GenBank 中23 种马圆线虫的线粒体cox1、nad1 和rrnL 基因序列两两进行比较,分析虫体间的基因相似性。

表2 序列分析和进化分析用圆线科线虫信息Table 2 Information of family strongylidae used in sequences and phylogenetic analysis
1.4 进化分析
以叶氏夏伯特线虫(Chabertia erschowi)为外群,cox1、nad1 和rrnL 基因序列串联,利用mafft7.0-37软件[12]进行比对,比对后利用在线软件Gblocks(http://molevol.cmima.csic.es/castresana/ Gblocks_server.html)[13]的默认值去除序列的基因间隔和模糊区域,用Clustal X 1.83 软件[14]进行格式的转换,采取BI 法,选取已有基因序列的23 种马圆线虫与研究中的6 种线虫(表2)构建进化树,探讨马圆线虫间的亲缘关系。
2 结果与分析
2.1 虫体鉴定
研究所6 种线虫的核糖体ITS 序列与GenBank中长伞杯冠线虫(KM085359.1)、冠状冠环线虫(JN786950.2)、细口杯环线虫(KP693432.1)、拉氏杯口线虫(KP693434.1)、麦氏副杯口线 虫(KP693435.1)和无齿圆形线虫(KP693438.1)的基因相似性分别为99.76%、99.07%、98.45%、99.88%、99.88%和99.36%。因此,研究所鉴定的虫体准确,可以进行后续试验。
2.2 序列分析
研究测得的6 种线虫的线粒体基因cox1 序列长度均为1 578 bp;无齿圆形线虫的nad1 核苷酸序列长度为879 bp,其余5 种线虫的核苷酸序列长度均为873 bp;6 种线虫的rnnL 序列全长均不同,分别为长伞杯冠线虫(976 bp)、冠状冠环线虫(974 bp)、细口杯环线虫(980 bp)、拉氏杯口线虫(978 bp)、麦氏副杯口线虫(973 bp)和无齿圆形线虫(959 bp)。cox1 基因与其他23 种圆线科线虫比较发现,除蝶状盅口线虫(Cyathostomum pateratum)的核苷酸序列长度为1 575 bp,氨基酸为524 个外,其余圆线科线虫的核苷酸序列长度和氨基酸数量均为1 578 bp 和525个。nad1 核苷酸序列长度除了圆形属线虫核苷酸序列稍长(876~879 bp)和氨基酸(291、292 bp)稍多外,其余线虫的核苷酸均为873 bp,氨基酸为290 个。而所有圆线科rnnL 序列长度则不规律,大小在959~983 bp之间(表2)。这6 种线虫的cox1 的A+T 含量在67.55%~69.71% 之间,AT 斜率在-0.28 与-0.24 之间;nad1 的A+T 含量在71.36%~73.83% 之间,AT 斜率在-0.33 与-0.29 之间;rnnL 的A+T 含量在80.88%~82.59%之间,AT 斜率在-0.04 与-0.03 之间。这些圆线科线虫的三种线粒体基因均呈AT 偏好。
将这6 种线虫的cox1、nad1 和rnnL 基因序列与GenBank 中其他23 种马圆线虫进行相似性分析发现,马圆线科线虫间的相似性分别为83.4%~97.9%,79.0%~98.5%和78.8%~98.9%。在cox1 基因分析中,无齿圆形线虫与马圆形线虫(Strongylus equinus)的相似性最高,为97.9%,锯齿三齿线虫(Triodontophorus serratus)和细口杯环线虫最低,为83.4%;在nad1基因中,无齿圆形线虫与马圆形线虫相似度最高,为97.3%,锯齿三齿线虫和普通圆形线虫(Strongylus vulgaris)相似性最低,为79.0%;在rnnL 中,无齿圆形线虫与马圆形线虫相似性最高,为98.9%,无齿圆形线虫与异齿杯口线虫(Poteriostomum imparidentatum)相似性最低,为79.5%。特别值得注意的是,短尾三齿线虫(Triodontophorus brevicauda)的cox1 基因与同亚科马圆形线虫的相似性为86.0%,而与盅口亚科的小唇片冠环线虫(Coronocyclus labratus)的相似性为88.1%;在nad1 的相似性分析中,短尾三齿线虫与普通圆形线虫的相似性为80.3%,而与盅口亚科的隐匿杯环线虫(Cylicocyclus insignis)为89.0%;对rnnL 的分析也是如此,短尾三齿线虫与普通圆形线虫的相似性为82.2%,与盅口亚科的隐匿杯环线虫为84.3%。比较分析还发现,三齿属线虫与盅口亚科的相似性总体要高于同亚科的圆线属。
2.3 进化分析
以串联的线粒体cox1、nad1 和rrnL 基因为标记,构建的系统进化树见图1。进化树形成了两大分支,一个分支为圆形属线虫,另一个分支包括盅口亚科所有线虫和三齿属线虫。在圆线属的分支中,无齿圆形线虫与马圆形线虫聚集一起,再与普通圆形线虫相聚,说明前两者关系更近。在另一大分支中,3 种三齿线虫形成一独立分支,而其他盅口亚科线虫形成另一分支。在盅口亚科分支中,所有的杯环属线虫均在一个分支中,虽然其中含有冠状冠环线虫,仍然认为其为单系;大唇片冠环线虫(Coronocyclus labiatus)和小唇片冠环线虫与两种杯冠线虫(长伞杯冠线虫和高氏杯冠线虫)及盅口属形成姊妹支;5 条微小杯冠线虫单独分支与长伞杯冠线虫和高氏杯冠线虫离的很远,因此推测杯冠属和冠环属应为复系;在盅口亚科线虫的最外侧,是麦氏副杯口线虫、拉氏杯口线虫和异齿杯口线虫形成一个小分支,但同属并没有聚集在一起。
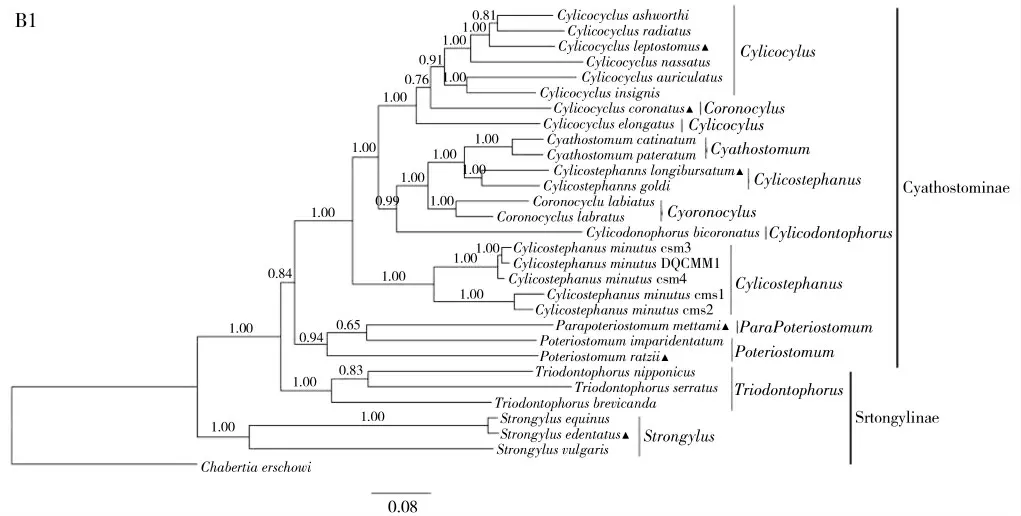
图1 基于线粒体cox1,nad1 和rnnL 串联序列通过BI 法构建的6 种线虫与其他圆线科线虫的进化树Fig.1 Phylogenetic tree of six worms with other strongylidae nematodes reconstructed by BI method based on combined mitochondrial genes cox1,nad1 and rnnL sequences
3 讨论
传统的马圆线虫间的亲缘关系研究以形态学为主,同属间的关系自然要近于不同属间,但在分类过程中可存在一些人为因素。随着科技的发展,分子生物学研究更具科学性,基于基因序列信息从分子层面反应物种进化过程中的异同,从而尽可能完整的还原物种的进化过程,体现它们间的亲缘关系[15-16]。线粒体基因组及其单独基因,特别是一些保守的基因,如cox1、nad1 在寄生虫分类学、种群遗传学和系统学研究中有着广泛的应用[8-9,16]。也有许多学者利用串联的线粒体基因对寄生虫进行遗传和分类探究,Wang 等[17]通过对来自黑龙江、吉林、陕西和云南四省的40 条羊仰口线虫(Bunostomum trigonocep-halum)的线粒体cox1,cytb,nad1 和nad5 基因序列进行序列分析和进化分析,结果表明不同地区的羊仰口线虫基因间存在着较低的种内变异,没有明显的地理区分;Lavikainen 等[18]利用串联的线粒体基因cox1 和nad1 基因序列对来自加拿大、土耳其、俄罗斯等国的Taenia crassiceps,Taenia hydatigena,Taenia martis,Taenia ovis krabbei 等绦虫进行进化分析,结果与以前采用单独基因的结果一致;Duan 等[19]对来自黑龙江、内蒙古的短尾三齿线虫(Triodontophorus brevicauda)和日本三齿线虫(Triodontophorus nipponicus)的线粒体cox1、cytb 和nad5 基因序列进行遗传变异分析,并通过三个序列串联进行进化分析,证明这些基因是序列分析和进化分析的良好基因标记。
研究获得了长伞杯冠线虫、冠状冠环线虫、细口杯环线虫、拉氏杯口线虫、麦氏副杯口线虫和无齿圆形线虫的线粒体基因cox1、nad1 和rrnL 序列。这6种线虫的三段线粒体基因序列与其他23 种圆线科线虫相比,cox1 和nad1 基因序列长度基本一致(蝶状盅口线虫、无齿圆形线虫、马圆形线虫和普通圆形线虫除外),而rrnL 在长度上差异较大,这与典型后生动物线粒体cox1、nad1 和rrnL 基因的长度特点一致[9-10,20]。研究6 种线虫的cox1、nad1 和rrnL基因序列的A+T 含量与目前已经被鉴定的大多数线虫相似[9-10],而A+T 含量的百分比明显高于吸虫,如华支睾吸虫(Clonorchis sinensis)(cox1:59.04%;rrnL:59.12%;nad1:62.13%)[21]。这6 种线虫rrnL 基因的A+T 含量的百分比大于cox1 和nad1 基因,类似的现象也出现在前人研究的其他线虫线粒体基因中,并且AT 斜率也与其他圆线科线虫相似[18,21-22]。在这三段不同基因的相似性比较分析中,无齿圆形线虫总是与马圆形线虫的相似性最高。麦氏副杯口线虫,拉氏杯口线虫和异齿杯口线虫与其他线虫的相似性均不高,这与Gao 等[9,23]的研究结果一致。更为重要的是,研究分析发现,cox1 和rrnL 基因序列比较分析中,29 种虫体的相似性虽然有高有低,但总体来看,三齿属线虫与同亚科圆线虫间的相似性要低于盅口亚科,而nad1基因比较中,三齿属线虫与圆线虫间的相似性均低于或等于盅口亚科。
研究基于线粒体cox1、nad1 和rrnL 基因序列构建了马圆线科线虫的进化树,对虫体间的亲缘关系进行探讨。在盅口亚科中,冠状冠环线虫与杯环属的线虫聚集在一起,与杯环属关系更近,研究结果与Zhang 等[24]以核糖体大亚基D3+区的序列信息构建的21 种马的圆线虫进化树结果相似,其结果也显示冠状冠环线虫与杯环属的线虫在同一分支内,因此冠状冠环线虫确实是一个比较特殊的虫体,也进一步证明冠环属为复系。5 条微小杯冠线虫(已被证明是复合种)单独分支,而长伞杯冠线虫和高氏杯冠线虫却聚集在一起,两组间距离却很远,这与Gao 等[10]利用线粒体全基因组和Huang 等[25]基于核糖体ITS序列探究30 种马圆线虫的进化关系分析的结果一致,因此支持微小杯冠线虫是复杂种的观点,认为杯冠属为复系。麦氏副杯口线虫和异齿杯口线虫先聚集,再与拉氏杯口线虫形成一小分支,研究结果与Tombak 等[26]利用ITS 基因序列结果不同,可能是他的分析中还有真臂副杯口线虫(Parapoteriostomum euproctus)的原因,因此更多的圆线虫相关基因被扩增应用十分重要。同时认为,副杯口属线虫、杯口属线虫与其他盅口亚科线虫关系更远。研究发现,三种三齿线虫并没有与同亚科的圆线属聚集在一起,而是与盅口亚科虫体在一起,研究结果与Gao 等[9]基于线粒体全基因组研究结果相似,因此研究结果也支持三齿属线虫属于盅口亚科的假说。
研究首次扩增了长伞杯冠线虫、冠状冠环线虫、细口杯环线虫、拉氏杯口线虫、麦氏副杯口线虫和无齿圆形线虫的线粒体基因cox1、nad1 和rrnL 的序列,并通过序列分析和进化分析探讨了马圆线科线虫间的亲缘关系,为马圆线虫的分类学、群体遗传学和系统学研究提供基础资料。
猜你喜欢
科普童话·百科探秘(2023年5期)2023-06-19 04:18:30
动漫界·幼教365(大班)(2021年1期)2021-04-06 04:15:21
武夷科学(2019年2期)2019-12-20 08:39:16
奥秘(2018年9期)2018-09-25 05:41:48
武夷科学(2017年0期)2017-03-10 10:03:31
浙江理工大学学报(自然科学版)(2016年5期)2016-09-15 05:49:11
初中生世界·七年级(2015年10期)2015-09-10 17:31:33
红蜻蜓·低年级(2015年6期)2015-06-15 16:00:20
作文周刊·小学三年级版(2009年16期)2009-07-27 09:24:14
意林(2005年1期)2005-05-30 08:53:56
