
乙酰乙酸通过抑制Nrf2 诱导奶牛乳腺上皮细胞线粒体损伤
2024-04-01 05:27宋倩高爽董昊徐闯孙旭东
黑龙江八一农垦大学学报 2024年1期
宋倩,高爽,董昊,徐闯,孙旭东
(黑龙江八一农垦大学动物科技学院,大庆 163319)
围产期是奶牛关键生理阶段,此时奶牛机体由于泌乳所需能量急剧增加的同时,干物质采食量(Dry matter intake,DMI)降低[1]。母牛难以从食物中获取足够的能量,会导致机体进入能量负平衡(Negative energy balance,NEB)状态[2]。机体为了适应NEB,将会进行脂肪动员,从而产生大量的脂肪酸以此满足机体能量所需。过量游离脂肪酸在肝脏内不完全氧化产生酮体,酮体过度累积导致奶牛酮病的发生,致使产奶量下降。奶牛乳腺上皮细胞是奶牛乳腺中合成以及泌乳的基本功能单位。奶牛乳腺上皮细胞功能紊乱导致乳腺组织功能障碍,泌乳量下降。研究发现,线粒体损伤导致奶牛乳腺上皮细胞功能异常,产奶量降低[3]。因此,阐明酮病奶牛乳腺上皮细胞损伤的分子机制将为我国养殖业提高奶牛产奶量提供帮助。
线粒体是产生能量的主要细胞器,为细胞提供所需能量[4]。因线粒体DNA(Mitochondrial DNA,mtDNA)是裸露的,并与呼吸链和富含脂质的线粒体膜紧密相连,使其较核DNA 对氧化损伤敏感性更高。线粒体DNA 异常引起线粒体功能障碍[5]。当线粒体功能障碍时,导致ROS 生成异常,打破ROS稳态,释放的大量ROS 导致线粒体膜电位下降,致使线粒体损伤[6]。研究发现临床酮病奶牛乳腺中存在线粒体功能障碍和氧化应激[7]。乙酰乙酸(Acetoacetic acid,ACAC)是奶牛体内酮体主要成分之一。但ACAC 对奶牛乳腺上皮细胞线粒体功能的影响尚不清楚。
核转录因子NF-E2 相关因子2(Nuclear factor E2-reated factor 2,Nrf2)是细胞氧化还原稳态的主要调节因子[8]。在非反刍动物中Nrf2 通过激活抗氧化基因保护细胞免受氧化应激[9]。除调节氧化还原的功能以外,Nrf2 也是重要的线粒体调节分子。Nrf2 可以通过直接和间接的方式调控过氧化物酶体增殖体激活受体γ-γ 活化剂1-α(Peroxlsome proliferator-activated receptor-γ coactlvator-1α,PGC-1α)、线粒体转录因子A(Mitochondrial transcription factor A,TFAM)、核呼吸因子-1(Nuclear respiratory factor-1,Nrf1)等线粒体生物合成的关键转录因子,促进抗氧化细胞的保护反应,使线粒体生物合成增加。研究发现在神经退行性疾病中,PGC-1α-Nrf-TFAM 途径的激活导致mtDNA 和蛋白质的合成以及生成新线粒体[10]。以上研究说明Nrf2 可以调节线粒体功能,但尚不清楚Nrf2 是否参与ACAC 对奶牛乳腺上皮细胞线粒体损伤的影响。
因此,试验在奶牛乳腺上皮细胞系MAC-T 添加不同浓度的ACAC 进行处理,来诱导线粒体损伤;并添加10 μmol·L-1的SFN,检测PGC-1α、TFAM、Nrf1与COⅠ-Ⅴ的基因表达情况,并检测ROS 和mtDNA 的含量及线粒体膜电位和ATP 含量的变化,旨在探讨ACAC 导致奶牛乳腺上皮细胞线粒体损伤的调控机制,为防治酮病奶牛代谢异常导致的乳腺组织线粒体损伤提供试验依据。
1 材料方法
1.1 主要试剂及仪器
DMEM-F12 培养基(Hyclone 公司);胎牛血清(CLARK 公司);商业ATP 测定试剂盒(南京建成生物工程研究所);Trizol 和RNA 反转录试剂盒购自TaKaRa;SYBR green plus(Roche,Norwalk,CT);722N分光光度计(上海精密仪器有限公司);荧光定量PCR仪(7500 Real-Time PCR 系统Applied Biosystems)。
1.2 细胞培养及处理
1.3 检测指标
1.3.1 线粒体ROS 的测定
将MAC-T 细胞用2 μg·mL-1Mito-roGFP 质粒(Addgene 质粒#49437)瞬时转染48 h。随后,通过流式细胞术分析线粒体ROS。通过计算405 nm 与488 nm荧光信号强度比值,分析线粒体ROS 含量。
1.3.2 ATP 含量的测定
使用商业ATP 检测试剂盒。在95 ℃,用500 μL去离子水将MAC-T 细胞匀浆10 min 后,在沸水中孵化与基质混合1 min,与底物溶液和催化剂混合,37 ℃孵化30 min,加入沉淀剂到混合物中,室温1 500×g 离心5 min,上清液与显色液室温孵育2 min后,室温下加入停止液5 min。吸光度在636 nm 处测定,将ATP 含量归一化为蛋白质浓度。根据制造商的说明,使用试剂盒(P1511;Applygen Technologies Inc.)检测总蛋白浓度。
1.3.3 RNA 分离和实时定量PCR(qPCR)
细胞在六孔板中处理结束后,按照每孔1 mL 的量加入Trizol,用移液枪吹打数次,并收集细胞于无RNA 酶的EP 管中。按照加入氯仿抽提—离心—加入异丙醇沉淀—离心—酒精洗涤—离心的步骤提取细胞总RNA。根据RNA 浓度,将1 μg RNA 样品使用试剂盒反转录为cDNA(RR047A,TaKaRa Biotechnology)。反应总体系(20 μL):酶10 μL;上下游引物各1 μL;DEPC 水6 μL;DNA 模板2 μL;反应程序见表1。使用SYBR green plus 试剂盒在7500 Real-Time PCR 系统检查靶基因的表达量;在Excel 软件中用2-ΔΔCT 方法计算目的基因的表达水平。引物合成依据Sun 等[12]研究中的引物合成方法,所用全部引物设计与合成来源于生工生物工程(上海)股份有限公司(见表2)。

表1 Real-time PCR 反应程序Table 1 Real-time PCR reaction program

表2 Real-time PCR 分析所用引物Table 2 Gene-specific primers used for Real-time PCR
1.3.4 线粒体膜电位检测
吸出每孔的培养液,PBS 洗涤细胞1 次,加入1 mL 培养液。加入1 mL JC-1 染色工作液,充分混匀,37 ℃孵育20 min。孵育期间按1 mL JC-1 染色缓冲液(5x)加入4 mL 蒸馏水比例,配制JC-1 染色缓冲液(1x),放置冰浴。37 ℃孵育结束后,吸去上清液,用JC-1 染色缓冲液洗涤2 次(冰浴),荧光显微镜下观察。
响应快捷化:①对当班发生的变化情况必须详细记录发生时间、地点、影响范围,按变化性质与要求及时汇报,立即启动响应程序;②对上一班延续的变化项目,要详细跟踪落实变化进展情况,确保闭合管理;③发生瓦斯等有害气体报警或超限时,必须在3分钟之内汇报上级部门并及时落实事故原因;④对所有变化环节响应,全部规范时间标准,要求所有响应步骤全部在规定时间内完成。
1.3.5 线粒体DNA 检测
采用凝胶电泳法获得DNA 完整数据。采用I2S rRNA 和亲环蛋白A 以及PrimeStar Max DNA 聚合酶(RO45A:TaKaRa 生物技术有限公司)扩增mtDNA 和核DNA(nDNA)。评估mtDNA 产物的相对数量。随后,用DNA 凝胶试剂盒(DOOS6;Beyotime 生物技术研究所)纯化pcr 产物。用阿伏伽德罗常数和模板DNA 的平均分子量计算每微升mtDNA 或nDNA 的拷贝数。
阿伏伽德罗常数是6.02×1023,C=浓度(ng·μL-1),Mwt=碱基对×660。根据拷贝数,将PCR 产物稀释10 次,从108倍到103copics·μL-1。采用SYBR Green定量RT-PCR 试剂盒(TaKaRa 生物技术有限公司),使用Applied 生物系统公司的7500 Real-Time PCR系统进行定量逆转录PCR(qRI-PCR)。采用循环阈值和浓度自然对数建立标准曲线,计算mtDNA 或nDNA 的拷贝数。mtDNA 的相对拷贝数换算为nDNA的相对拷贝数。
1.4 统计学分析
采用IBM SPSS Statistics 26 软件对各组试验数据进行单因素方差分析,各组间的显著性差异用字母表示(P<0.05),数据表示形式为平均数±标准误。
2 结果与分析
2.1 ACAC 对线粒体活性氧和线粒体膜电位的影响
与0 mmol·L-1ACAC 对照组相比,1.2 和1.8 mmol·L-1ACAC 处理组的MAC-T 细胞中线粒体ROS 含量显著升高(图1(A),P<0.05),而与对照组相比,1.2 和1.8 mmol·L-1ACAC 组线粒体膜电位显著降低(图1(B),P<0.05)。
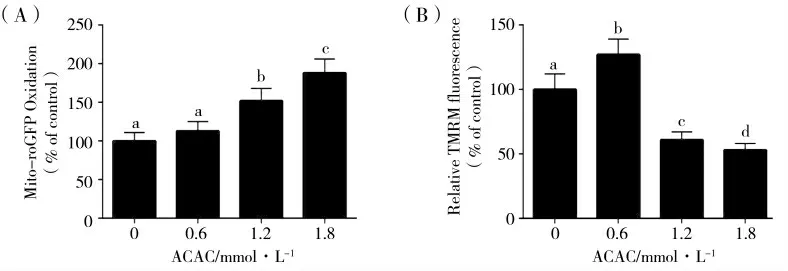
图1 ACAC 对ROS 和线粒体膜电位的影响Fig.1 Effects of ACAC on ROS and mitochondrial membrane potential
2.2 ACAC 对线粒体呼吸链和ATP 含量的影响
与对照组0 mmol·L-1ACAC 相比,1.2 和1.8 mmol·L-1ACAC 处理组COⅠ、COⅡ、COⅢ、COⅣ以及COⅤ的mRNA 表达水平显著降低(图2(A)(B)(C)(D)和(E),P<0.05)。与对照组相比,1.2 和1.8 mmol·L-1ACAC 处理组的ATP 含量显著降低(图2(F),P<0.05)。

图2 ACAC 对线粒体呼吸链和ATP 含量的影响Fig.2 Effects of ACAC on mitochondrial respiratory chain and ATP content
2.3 ACAC 对线粒体DNA 和NFE2L2 基因表达水平的影响
与对照组相比,1.2 和1.8 mmol·L-1ACAC 处理组NFE2L2、PGC-1α、Nrf1 以及TFAM 的mRNA 表达量显著降低(图3(A)(B)(C)和(D),P<0.05)。与对照组相比,1.2 和1.8 mmol·L-1ACAC 处理组mtDNA 含量变化显著降低(图3(E),P<0.05)。

图3 ACAC 对线粒体含量和NFE2L2 基因表达水平的影响Fig.3 Effects of ACAC on mitochondrial content and NFE2L2 gene expression
2.4 SFN 预处理缓解了ACAC 诱导的线粒体含量下调
与DMSO 组相比,ACAC+DMSO 处理组Nrf2 的mRNA 表达水平显著降低(图4(A),P<0.05)。与ACAC+DMSO 处理组相比,ACAC+SFN 处理组Nrf2 的mRNA 表达水平显著升高(图4(A),P<0.05)。与DMSO 组相比,ACAC+DMSO 处理组PGC-1α、Nrf1以及TFAM 的mRNA 表达量显著降低(图4(B)(C)和(D),P<0.05)。与ACAC+DMSO 处理组相比,ACAC+SFN 处理组的PGC-1α、Nrf1 以 及TFAM 的mRNA 表达量显著升高(图4(B)(C)和(D),P<0.05)。与DMSO 组相比,ACAC+DMSO 处理组的mtDNA 含量显著降低(图4(E),P<0.05)。与ACAC+DMSO 组相比,ACAC+SFN 组mtDNA 含量显著升高(图4(E),P<0.05)。
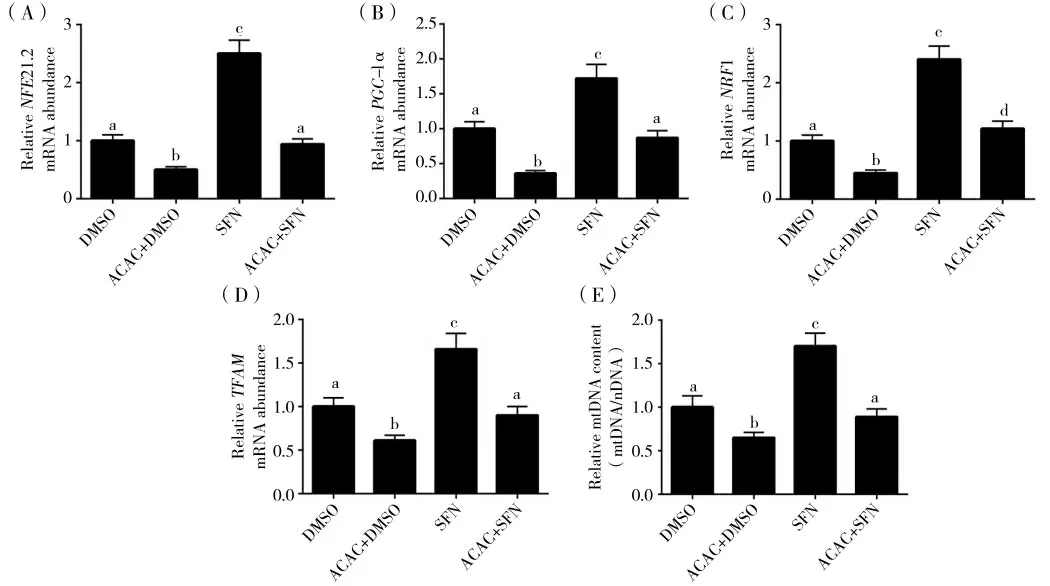
图4 SFN 缓解ACAC 诱导Nrf2 表达下降及线粒体含量下调Fig.4 SFN alleviates ACAC-induced Nrf2 expression and mitochondrial content down-regulation
2.5 SFN 预处理缓解了ACAC 诱导的线粒体损伤
与DMSO 组相比,ACAC+DMSO 处理组MAC-T细胞中的ROS 含量显著升高(图5(A),P<0.05),线粒体膜电位显著降低(图5(B),P<0.05)。与ACAC+DMSO 处理组相比,ACAC+SFN 处理组的MAC-T 细胞中的ROS 的含量显著降低(图5(A),P<0.05),而线粒体膜电位显著升高(图5(B),P<0.05)。与DMSO组相比,ACAC+DMSO 处理组的COⅠ、COⅡ、COⅢ、COⅣ和COⅤ的mRNA 表达量显著降低(图5(C),P<0.05)。与ACAC+DMSO 组相比,ACAC+SFN 预处理组COⅠ、COⅡ、COⅢ、COⅣ和COⅤ的mRNA 表达量显著增加(图5(C),P<0.05)。与DMSO 组相比,ACAC+DMSO 处理组的ATP 含量显著降低(图5(D),P<0.05)。与ACAC+DMSO 处理组相比,ACAC+SFN 处理组的ATP 含量显著增加(图5(D),P<0.05)。

图5 SFN 缓解ACAC 诱导线粒体损伤Fig.5 SFN alleviates ACAC-induced mitochondrial damage
3 讨论
奶牛酮病发生的主要诱因之一是NEB[11]。严重的能量负平衡会导致奶牛代谢紊乱,诱导奶牛体内产生大量的游离脂肪酸和酮体。研究发现,代谢紊乱可以诱发酮病奶牛乳腺组织线粒体损伤,产生大量的ROS,并导致产奶量降低[12]。因此,阐明代谢应激导致奶牛乳腺上皮细胞线粒体损伤的分子机制,对防治酮病奶牛乳腺上皮细胞功能紊乱至关重要。研究结果显示,高浓度ACAC 通过抑制Nrf2 表达,诱导奶牛乳腺上皮细胞线粒体损伤,导致线粒体ROS 蓄积,因此推测Nrf2 可能是防止代谢应激导致酮病奶牛乳腺组织线粒体损伤的治疗靶点。
线粒体是由核基因组和线粒体基因组共同调控的细胞内双层膜结构的动态细细胞器[13]。线粒体生物发生异常,导致线粒体功能障碍打破细胞稳态。线粒体是产生ROS 的主要场所。因线粒体DNA 裸露导致线粒体更易受到ROS 攻击[14],引起线粒体功能紊乱,致使细胞氧化损伤。已有研究证明,ACAC 诱导奶牛肝细胞氧化应激[15]。研究结果表明,高浓度的ACAC处理,导致奶牛乳腺上皮细胞中线粒体ROS 含量升高。以上结果说明代谢应激导致奶牛乳腺上皮细胞氧化损伤。细胞损伤时ROS 增加,导致线粒体膜电位降低,导致线粒体功能障碍。研究证实,高浓度NEFA处理奶牛肝细胞ROS 含量增加,导致线粒体膜电位降低,致使线粒体损伤[16]。研究结果发现高浓度的ACAC 处理,导致奶牛乳腺上皮细胞中线粒体膜电位降低。这些结果表明,ACAC 通过诱导线粒体ROS 增加,导致奶牛乳腺上皮细胞中线粒体膜电位降低,致使奶牛乳腺上皮细胞功能障碍。线粒体是产生能量的主要场所,为细胞提供90%的能量[17]。线粒体主要通过细胞器内膜中的5 个多亚基复合物(COⅠ-Ⅴ)进行氧化磷酸化合成ATP[18]。前人研究显示,提高小鼠体内ATP 合成底物使用效率与功能活性,使得乳汁合成速率提高[19]。研究中高浓度的ACAC 导致COⅠ-Ⅴ的基因表达降低,并且ATP 含量降低。Song 等[7]研究表明,酮病奶牛诱乳腺组织线粒体活性氧增加和ATP 降低,导致线粒体功能障碍,产奶量下降。因此推测,代谢应激可能通过导致线粒体损伤,减少能量产生,抑制乳腺上皮细胞的泌乳能力。
Nrf2 是细胞氧化还原稳态的主要调节因子,在线粒体生物发生也发挥着重要作用[20]。研究发现,Nrf2激活有助于在应激条件下保持线粒体形态和功能[21]。线粒体生物发生涉及线粒体和核转录因子的协调调节。PGC-1α 是一种关键的转录共激活因子。TFAM负责mtDNA 的转录和复制。Nrf1 是一种核转录因子,控制涉及线粒体呼吸功能的靶基因。在非反刍动物中,PGC-1α 的增加,诱导Nrf1 和Nrf2 的转录,导致TFAM 激活,产生TFAM 蛋白质因子。TFAM 蛋白质因子转运进入线粒体内,与线粒体DNA 结合,刺激线粒体生物发生。试验结果显示,与对照组相比,高浓度ACAC 处理组PGC-1α、TFAM、Nrf1 以及Nrf2的基因表达量与mtDNA 含量显著降低。在非反刍动物中的研究表明,抑制PGC-1α、Nrf1、Nrf2 和TFAM的表达,导致线粒体生物合成的减少以及细胞代谢的改变[22]。因此,以上结果都说明ACAC 通过抑制Nrf2 表达,导致的乳腺上皮细胞线粒体功能障碍。
萝卜硫素(Sulforaphane,SFN)是一种异硫氰酸酯植物化学物质,具有抗氧化作用[23]。SFN 作为Nrf2 的激活剂,广泛应用到线粒体功能障碍性疾病治疗中[24]。研究发现SFN 可以通过激活Nrf2,促进线粒体生物发生、降低线粒体ROS 产生,缓解线粒体功能紊乱[25]。在研究中,SFN 预处理显著增加奶牛乳腺上皮细胞中Nrf2 的基因表达水平,缓解ACAC 对线粒体生物发生关键分子PGC-1α、TFAM 和Nrf1 mRNA 表达及mtDNA 含量的抑制作用。此外,SFN 预处理缓解了ACAC 诱导的线粒体ROS 含量的升高和线粒体膜电位升降低、COⅠ-Ⅴ的基因表达水平降低以及ATP含量降低。因此,以上结果表明通过SFN 激活Nrf2,能有效缓解高浓度ACAC 诱导的奶牛乳腺上皮细胞线粒体损伤。
4 结论
综上所述,数据表明ACAC 通过抑制Nrf2 的表达水平,抑制奶牛乳腺上皮细胞线粒体生物发生、降低能量生成,诱导ROS 蓄积。而SFN 通过增加Nrf2的基因表达,缓解ACAC 诱导的线粒体损伤。研究证实,过量的ACAC 可以抑制Nrf2,并诱导奶牛乳腺上皮细胞MAC-T 线粒体功能障碍。
猜你喜欢
今日农业(2022年3期)2022-11-16
安家(校外教育)(2022年6期)2022-01-03
世界科学技术-中医药现代化(2021年7期)2021-11-04
今日农业(2021年10期)2021-07-28
海洋通报(2021年1期)2021-07-23
生物学通报(2021年4期)2021-03-16
小天使·一年级语数英综合(2019年12期)2019-01-13
小天使·三年级语数英综合(2014年9期)2014-09-12
中国药理学通报(2014年2期)2014-05-09
癌变·畸变·突变(2014年1期)2014-03-01
