
穗分化期高温对水稻生长发育的影响及缓解措施研究进展
2024-03-23 09:21丁文金何招亮戴彬凤钟列权张海鹏
浙江农业学报 2024年2期
郑 涵,丁文金,何招亮,侯 凡,戴彬凤,钟列权,张海鹏,*,杨 勇
(1.安徽农业大学 农学院,安徽 合肥 230036; 2.浙江省农业科学院 病毒学与生物技术研究所,农产品质量安全危害因子与风险防控国家重点实验室,农业农村部和浙江省植保生物技术重点实验室,浙江 杭州 310021; 3.浙江勿忘农种业股份有限公司,浙江 杭州 311200; 4.台州市农业生态保护与质量安全中心,浙江 台州 318000)
水稻(OryzasativaL.)是世界上最重要的粮食作物之一,自20世纪70年代以来我国长江中下游稻区单季稻种植面积逐年增加,该地区作为我国重要的水稻产地,对保障我国水稻产量有重要意义[1-2]。随着工业化程度的不断推进,温室气体的排放量逐年增加,高温天气出现的频率和强度也呈现出上升趋势[3]。长江中下游地区因其特殊的地理和环境因素,高温和伏旱天气在该地区夏季频繁交替出现,而夏季又恰好是单季稻穗分化的关键时期,高频率出现的高温天气已对该地区水稻的安全生产产生了极大威胁,不同水稻品种和生育期的不同对于穗分化期高温的耐受性不同[4-6]。通常情况下穗分化期高温会通过影响水稻幼穗形成、花粉发育、调控代谢生理指标等过程进而导致水稻减产。研究发现穗分化期高温会显著降低水稻每穗粒数与千粒重并因此导致水稻严重减产[6]。本文从穗分化期高温导致水稻幼穗发育不良、花粉育性降低、产量和品质下降、生理代谢紊乱等方面综述了穗分化期高温对水稻生长发育的影响并提出相应的缓解措施和前景展望。以期为开展水稻逆境生理研究以及缓解措施的合理选择提供依据。
1 穗分化期高温对水稻生长发育的影响
穗分化期是水稻生长过程中非常重要的一个时期,一般情况下,根据水稻自身生长情况以及生产研究的实际又将穗分化期分为8个时期[7]。又根据水稻幼穗发育情况的不同,前4个时期被称为幼穗形成期,后4个时期则被称为孕穗期,高温作用时间的不同会对水稻生长发育造成不同影响。以下将重点归纳穗分化期高温对幼穗形成与发育、花药的结构和花粉发育以及对水稻产量的影响3个方面。
1.1 对幼穗形成与发育的影响
幼穗形成与发育是水稻产量构成的基础。水稻作为喜温植物,需要在温度较高的环境下才可进行正常的生长与发育,通常情况下环境温度保持在26.0~33.1 ℃最有利于幼穗形成,低于或者高于此温度范围皆会造成水稻幼穗形成受阻[8-9]。研究显示,高温导致水稻穗分化期减数分裂的颖花和枝梗显著退化,并且不同品种穗间位存在差异,造成水稻幼穗的不正常发育,使水稻幼穗分化畸形[10]。通常情况下,幼穗形成需要水稻提供大量的碳水化合物,因此碳水化合物合成与转运的减少是影响幼穗发育的主要原因[11-12]。陈燕华等[13]研究发现,穗分化期高温胁迫下水稻光合作用并未受到显著影响,水稻幼穗发育受到制约是因为糖转运受阻造成的,高温胁迫显著下调了蔗糖转运基因OsSUT1、OsSUT2、OsSUT4,上调了细胞分裂素氧化酶基因OsCKX5、OsCKX9的表达,进而降低了幼穗中非结构性碳水化合物(NSC)的积累并最终抑制了水稻颖花的分化并加剧了颖花退化。Chen等[14]的研究进一步证实了这一观点,即在穗分化期高温胁迫下水稻干物质的积累并未受到影响,高温通过降低细胞分裂素(CTK)、赤霉素(GA)、吲哚乙酸(IAA)等激素含量、抗氧化酶活性以及糖酵解和三羧酸循环相关酶活性,提高脱落酸(ABA)含量、CTK氧化分解基因OsLOGL2和OsLOGL3的表达量以及膜脂过氧化物含量,最终降低了水稻幼穗对于蔗糖的利用效力以及NSC的积累量。但是对于这一现象,有研究者却发现穗分化期高温不仅会影响到“流”(蔗糖转运),同时“源”(叶片光合能力)也会受到影响和限制,并因此造成水稻幼穗发育不良[15]。穗分化后4个时期的高温同样会对水稻幼穗形成造成影响,后4个时期因为颖花分化已基本完成,因此在这一时期,高温主要通过提高水稻颖花退化率进而影响水稻幼穗的形成[16]。王亚梁等[17]在水稻倒一叶期进行40 ℃高温胁迫后发现水稻颖花退化显著,并且随着高温时间的延长,颖花退化情况越明显。综上所述,高温下水稻幼穗干物质积累量减少是造成幼穗发育不良的主要原因。
1.2 对花药结构和花粉发育的影响
1.2.1 对花药结构的影响
水稻花药是水稻颖花的重要组成部分之一,也是可育花粉形成的基础[18]。穗分化期高温下水稻花药结构受损严重、花粉发育受阻,其中绒毡层形态异常、花药表皮角质层结构以及花药壁加厚都是影响花药结构发育的重要原因[19]。水稻花药绒毡层是花药中供给花粉发育营养物质的重要器官,其上分泌的脂类物质对于花粉壁形成以及花粉的正常发育皆具有重要作用[20]。高温下,绒毡层结构可通过UPR(未折叠蛋白)反应来感知高温并促使内质网结构改变进而提高花药对高温的耐受性[21]。但是同时,高温也会通过增加花药内ABA含量、下调NADPH氧化酶调控基因OsPDIL1-1以及超氧化物歧化酶(SOD)调控基因Cu/Zn-SODa基因的表达、降低SOD、过氧化氢酶(CAT)等抗氧化酶活性导致水稻花药中的活性氧(ROS)含量激增进而阻碍花药绒毡层细胞的程序性死亡(PCD)过程,PCD过程的受阻则直接导致了花药绒毡层功能的缺失[22-25]。目前有研究发现bHLH转录因子、PHD-finger结构域蛋白质等转录因子与绒毡层的PCD过程密切相关[26-27]。但是目前的相关研究仍缺少高温下相关转录因子与绒毡层PCD相互关系的研究,未来可就此作为创新点展开深入研究。此外,作为花药重要组成部分的药隔维管束在水稻花器官的发育过程中起着营养物质传输通道的重要作用,张桂莲等[28]在对水稻进行高温胁迫后发现,水稻花药中的药隔维管束结构被破坏,阻碍了营养物质运输,造成花器管发育不良。
1.2.2 对花粉发育的影响
穗分化期高温影响水稻花药结构,进而导致水稻花粉发育过程中营养物质供给不足是致使水稻花粉发育不良、育性降低的主要原因之一。除此之外,高温也会影响水稻花粉细胞的分裂过程从而造成花粉不育[29]。孕穗期是水稻花粉形成的重要时期,花粉母细胞首先经过一次减数分裂形成小孢子,小孢子在经过一次有丝分裂后形成生殖细胞,最后生殖细胞再经过一次有丝分裂才可形成具有育性的花粉细胞[30-31]。这时期的高温主要通过影响减数分裂以及有丝分裂的进程从而造成水稻花粉育性降低。例如,在水稻花粉母细胞进行第一次减数分裂期间,若此时遭遇高温则会影响减数分裂进程,致使同源染色体基因重组的概率增加,在减数分裂的后期同源染色体分裂失败,最终在形态上表现为形成异常小孢子[32-33]。同时Pan等[34]在分子层面对此进行了研究和分析,结果显示,穗分化期高温会通过影响调控多种锌指蛋白、多嘧啶束结合蛋白以及DEAD/DEAH盒RNA解旋酶等相关基因的表达进而影响水稻正常的减数分裂过程。此外,小孢子第一次有丝分裂是一次非极性的有丝分裂,通常情况下小孢子会分裂为一个生殖细胞以及一个营养细胞,若此时经历高温则会导致蔗糖转化酶(INV)活性降低,INV基因表达下调,淀粉积累异常并最终导致小孢子发育异常,分化异常[35-36]。综上所述,高温会通过影响水稻花粉细胞分裂过程进而造成水稻花粉发育异常甚至数量减少。
1.3 对水稻产量与品质形成的影响
水稻产量主要是由有效穗数、每穗粒数、结实率、千粒重4大因素构成的,穗分化期高温主要通过影响幼穗与花粉粒的形成发育进而影响产量。穗分化高温影响水稻颖花的分化与退化过程,导致水稻每穗粒数降低并最终影响产量[37]。同时,水稻花粉粒发育不良则会导致水稻结实率下降进而造成减产[38]。而关于水稻千粒重的变化,宋有金等[6]和陈燕华等[13]的试验则均显示水稻千粒重在穗分化期高温胁迫下均出现不同程度的降低。如表1显示穗分化期高温对水稻产量的影响。综上所述,根据目前的研究结果来看,穗分化期高温主要通过影响每穗粒数、结实率以及千粒重的方式来降低水稻产量。穗分化期高温也会对稻米品质造成影响,Mohammed等[39]研究发现,孕穗期夜间高温会降低籽粒的粒宽,提高垩白度,从而降低稻米品质。
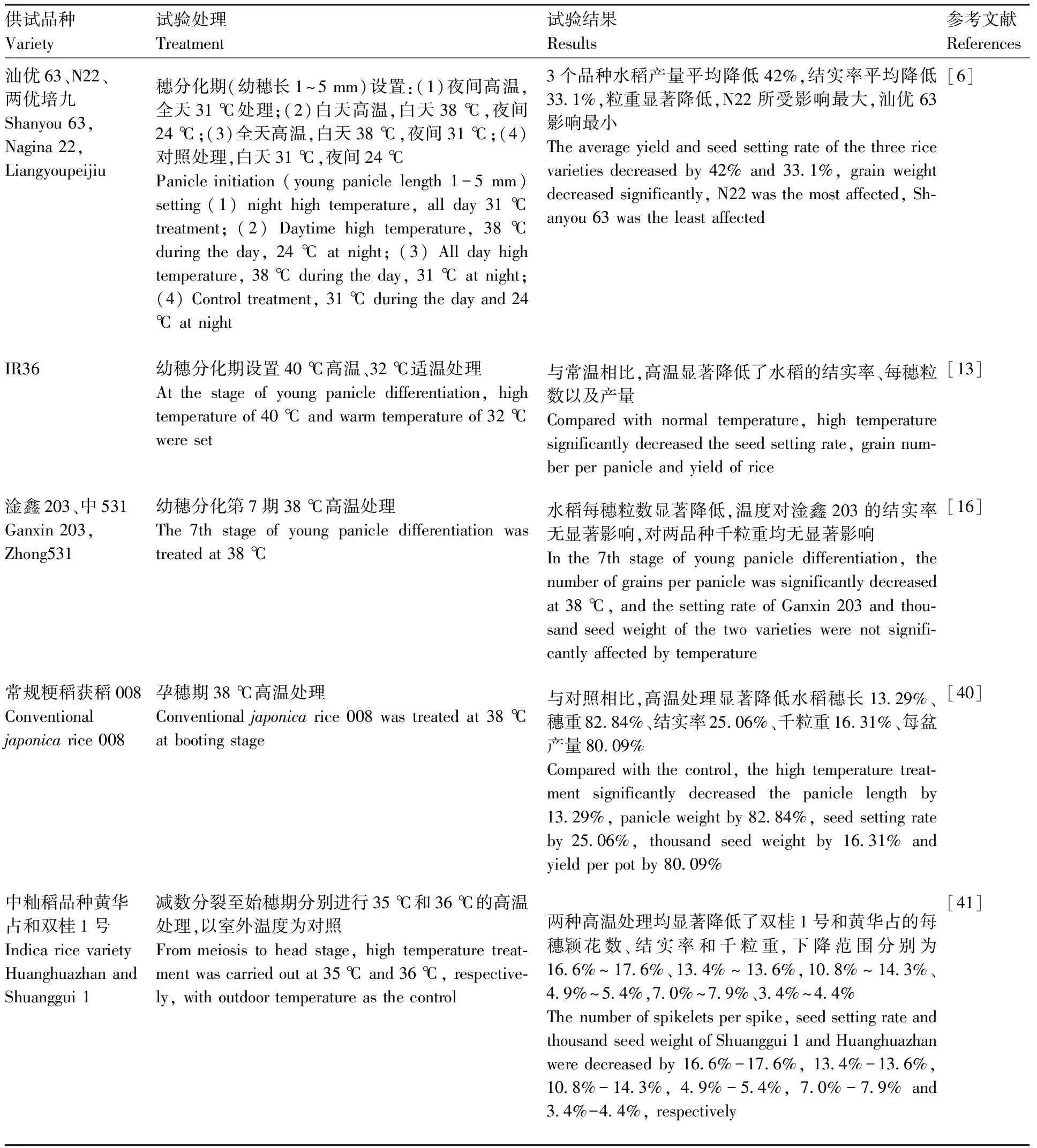
表1 穗分化期高温对水稻产量的影响
2 穗分化期高温对水稻生理特性的影响
穗分化期高温所引起的水稻生理变化是影响水稻生长发育的重要因素,其中光合特性、内源激素、抗氧化性的变化是其中最主要的几个原因。
2.1 对光合特性的影响
光合作用是植物进行生长发育和生理代谢的最重要环节,是水稻进行正常干物质同化和积累的基础[41-43]。Tang等[44]研究发现,穗分化期高温胁迫下水稻气孔导度(Gs)和蒸腾速率(Tr)以及光系统Ⅱ实际光化学效率(ΦPSII)显著下降。Gs、Tr以及ΦPSII是衡量水稻光合效率的几个重要指标。一般情况下水稻冠层温度与Gs和Tr关系密切,较低的冠层温度是保证颖花育性的基础,耐热性品种通常具有较高的Gs、Tr水平以保证对于高温的耐受性[44-45]。ΦPSII是关系水稻净光合速率(Pn)的重要指标,有研究发现穗分化期高温胁迫下,水稻叶片的Pn、SPAD、叶绿素含量、光系统Ⅱ最大光化学效率(Fv/Fm)、ΦPSⅡ、光化学猝灭系数(qP),以及非光化学猝灭系数(NPQ)均呈现不同程度的降低[46-47]。以上指标的变化显示出穗分化期高温胁迫下,水稻叶片的光合能力受到损伤,同化物合成以及固定能力减弱。唐汇春等[48]的研究显示,孕穗期高温胁迫下水稻Pn显著下降,Pn显著下降的同时伴随着水稻干物量积累以及转运量的减少,而干物质量积累与转运不足则直接导致了水稻产量的下降。又因为糖消耗的减少是导致水稻幼穗形成与发育的主要原因[37],所以据此可以推断出光合能力的减弱对于水稻穗分化期干物质积累以及幼穗发育有着较大关联。但是在本文前期的相关描述中,对于穗分化期高温对水稻光合的减弱程度以及干物质量积累还是转运二者谁是影响此时期水稻生长的最主要因素,目前研究仍存在着争议。Li等[49]研究发现,适度的高温条件下,水稻生长受到抑制的主要原因是呼吸作用加强而并不是光合能力减弱所导致的。结合前人的研究结果来看,供试品种本身对于高温的耐受性以及怎样的温度设置的确有可能是现今光合作用在穗分化期高温下作用机理存在差异的一个重要原因。
2.2 对内源激素的影响
内源激素含量变化是穗分化期高温影响水稻生长发育的又一个重要原因,IAA、CTK、GA、油菜素甾醇类化合物(BR)、ABA作为植物体内最主要的几类激素在此阶段表现出了不同的时空变化特征。IAA在花粉的发育过程中充当着重要作用,Tang等[50]研究发现,高温下IAA含量下降,水稻花粉育性降低,因此IAA含量的变化是导致水稻发育不良的主要原因。Cecchetti等[51]的研究结果显示,植物花药的发育后期中层IAA含量增加,若此时缺少对于IAA的感知能力,则会导致附近组织中的IAA含量积累加剧,最终引起花药发育不良、花粉失活。因此,IAA影响水稻花粉育性不仅在于含量变化,也在于空间上的变化。CTK是水稻幼穗发育过程中的关键激素,穗分化期高温会通过降低水稻幼穗中的CTK含量、抑制异戊烯基转移酶和细胞色素P450单氧化酶等CTK合成酶的活性、增强细胞分裂素氧化酶/脱氢酶的活性的方式降低幼穗中的CTK含量及其利用效率进而造成水稻幼穗发育不良[52-53]。GA和BR同样是两种参与水稻幼穗和花粉发育的重要激素,高温会降低这两种激素的含量从而导致幼穗与花粉的发育进程减缓[54-55]。穗分化期高温显著降低了内源BR含量,进而导致水稻幼穗中蔗糖转运基因OsSUT1、OsSUT2和OsSUT4的表达量下降,参与蔗糖水解、糖酵解和三羧酸循环的关键酶活性降低,最终导致幼穗发育中的糖分供给不足,发育不良[56]。事实上,IAA、CTK、GA以及BR通常在水稻穗分化过程中发挥着协同效应,但ABA却与以上激素之间呈现出拮抗作用。例如曹云英等[57]研究发现,水稻减数分裂期高温胁迫下,水稻幼穗中IAA、CTK等激素含量下降,ABA含量却呈现上升趋势。ABA作为一种逆境激素,在植物的应激反应中充当着重要角色,少量ABA对于植物抵御逆境有着积极作用,但大量ABA的积聚则会造成植物抗逆性减弱[58-59]。在水稻穗发育的过程中,高温诱导ABA在花药中大量积聚,造成ROS含量在花药中急速增加,影响绒毡层PCD过程,并最终致使花药结构被破坏,花粉发育不良[60]。各种激素在水稻穗分化期的生长过程中均充当着重要角色,并且在高温环境下,通常存在着几类激素间的协同或拮抗作用。
2.3 对氧化与抗氧化性的影响
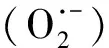
3 穗分化期高温的缓解措施
选用合理的缓解措施是水稻抵御高温危害、维持产量的最有效方法。现主要有调整栽培模式、喷施外源物质、选育耐热品种等措施可供选择。
3.1 调整栽培模式
调整合理的栽培模式是抵御水稻穗分化期高温胁迫的有效措施之一。穗分化期高温会通过影响水稻的冠层温度进而导致颖花育性降低。在实际的生产过程中,可以通过调整灌溉模式来降低土壤和水稻的冠层温度,保证水稻颖花育性[68]。同时合理施肥也是水稻穗分化期抵御高温的有效措施。Yang等[69]研究发现穗分化后期高温胁迫下,与低施氮相比,高氮肥处理显著提高了水稻的光合作用能力以及改善了水稻的生理内环境。因此,在实际的生产过程中,在大田栽培的模式下可以通过适当调整水肥管理和栽培模式的方法缓解穗分化期高温造成的水稻减产问题。
3.2 喷施外源物质
合理喷施外源物质是增强水稻抗逆性,提高水稻产量的重要措施。合理喷施外源物质对于水稻在穗分化期高温下的叶片光合作用、蔗糖转运与利用效率、抗氧化能力均有不同程度的提高[70-71]。例如穗分化期高温胁迫下外源喷施IAA可显著降低水稻体内的H2O2以及MDA含量、提高叶片叶绿素和脯氨酸含量进而提高水稻叶片的光合和抗氧化能力,降低水稻细胞在高温逆境下的膜脂过氧化程度[72]。陈燕华等[13]研究发现,穗分化期高温胁迫下BR有利于提高蔗糖的转运与利用效率以及水稻的抗氧化能力。同时,外源喷施水杨酸和褪黑素对于水稻缓解高温热害也具有显著的效果,可提高水稻的耐热性[73-74]。油菜素内酯可诱导植物激素相关合成基因的表达,增加内源BR、CTK、GA和ABA的含量,从而促进幼穗中蔗糖的分解和利用,增强超氧化物歧化酶、过氧化氢酶和过氧化物酶的活性,减少幼穗中H2O2和MDA的积累,可提高幼穗对蔗糖的利用[14]。水稻减数分裂期高温下,外源喷施ABA可显著提高水稻幼穗中可溶性糖、淀粉和NSC含量,增加热激蛋白(HSP24.1、HSP71.1)的相对表达,上调糖代谢和运输相关基因SUT、蔗糖合成酶(SUS)基因和INV基因的表达以及水稻幼穗的增强抗氧化能力[75]。因此,在实际的生产过程中可以通过合理选用外源喷施物质的方式缓解水稻穗分化期高温对水稻生长发育的损伤。
3.3 培育和选用耐热品种
耐热品种的选育是目前作物遗传育种专业研究的热点与难点。现代生物技术的发展为耐热水稻品种的选育提供了新思路。水稻穗分化期高温对水稻花药绒毡层结构影响较大,而在水稻绒毡层发育的过程中,MADS、bHLH和PHD-finger是几种控制绒毡层发育的重要的转录因子[76]。同时,bZIP、MYB/MYC、WRKY、AP2/EREBP和NAC等转录因子在水稻抗逆反应中也发挥着重要作用[77]。此外,在现今的生产过程中选用耐热性较强的水稻品种也是抵御穗分化高温的有效办法。例如,宋有金等[6]研究发现,汕优63在穗分化期、花期、灌浆期都表现出了较强的耐热性,而N22和两优培九均表现出了对于穗分化期高温的敏感性。因此,在实际生产中,种植户可以选用例如汕优63这样具有强耐热性的水稻品种,以此提高水稻产量。
4 总结与展望
穗分化期是决定水稻颖花数和结实率的关键时期。因此,夏季高温天气的频发与水稻最终的产量有着密切的关系。穗分化期高温对水稻生长发育的影响是复杂而多面的,首先对水稻的形态特征有所改变,使得水稻株高下降,叶鞘长度增加,柱头花粉数量降低,花药结构异常,籽粒外形改变等。其次,水稻的生理指标也有所变化,水稻气孔导度和蒸腾速率显著下降,高温胁迫下造成水稻叶片的叶绿素含量、净光合速率等指标不同程度下降,可溶性糖含量下降,细胞内源激素IAA、CTK、GA 等下降,ABA大量积累,活性氧含量增加,抗氧化酶活性降低等。通过对其代谢调控生理的研究表明,高温可影响水稻蔗糖转运蛋白基因OsSUT1、OsSUT2、OsSUT4等的表达,热激蛋白、蔗糖合成酶基因、糖代谢以及运转基因表达异常,而且高温导致幼穗内内源激素代谢紊乱,幼穗细胞分裂素含量显著下降,花粉内生长素含量减少,导致碳水化合物运转利用低下。最后,水稻的每穗粒数下降,结实率下降,千粒重下降,垩白度增加,影响稻米产量和品质。就目前的研究来看,学者普遍认为穗分化期高温导致水稻产量下降主要是由于碳水化合物供给和分配不平衡等原因导致的。针对这一现象,从不同角度解释了穗分化期高温导致水稻产量下降的原因并提出了相应的缓解措施(图1)。
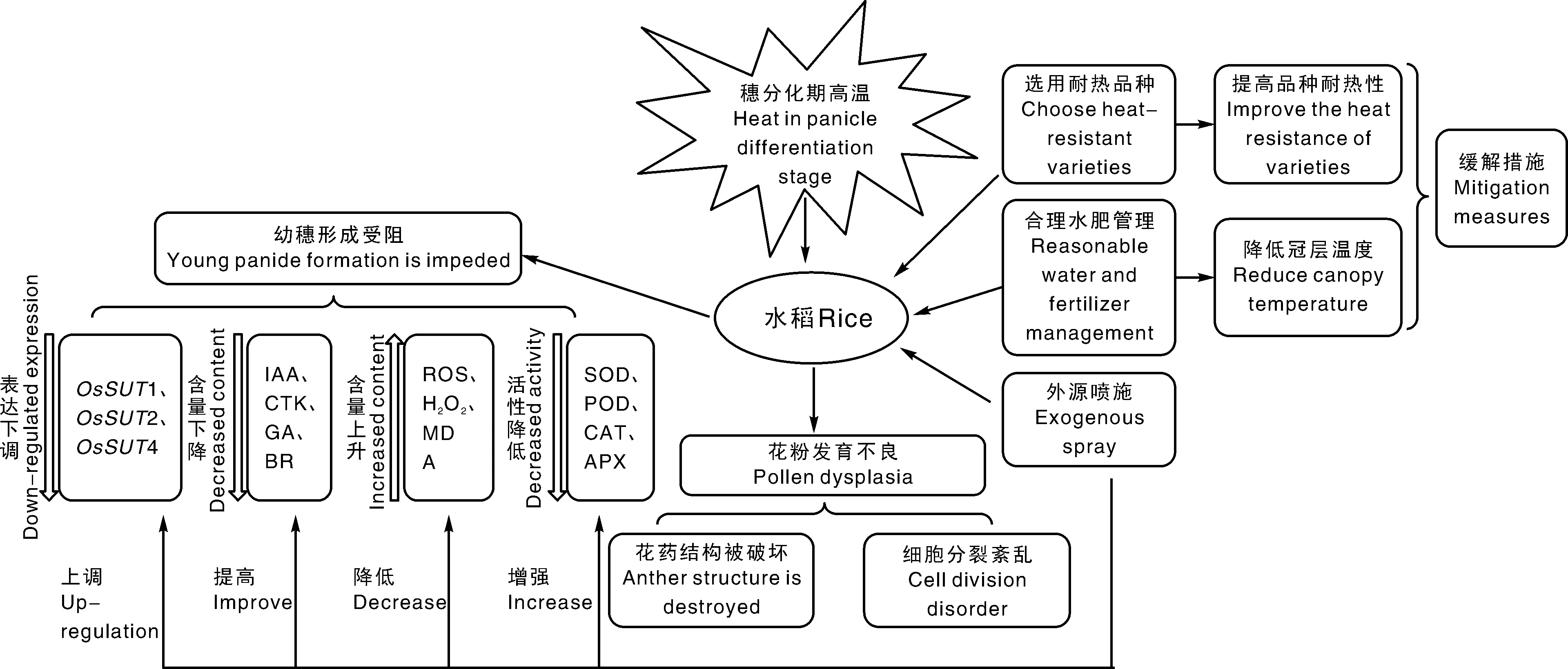
图1 穗分化期高温对水稻生长发育的影响及其缓解措施Fig.1 Effects of high temperature on rice growth and development during panicle differentiation stage and its mitigation measures
目前,穗分化期高温导致长江中下游稻区水稻幼穗发育不良,颖花育性降低,造成水稻大幅度减产的问题依然存在。结合国内外对穗分化期高温对水稻生长发育以及生理特性的研究,笔者认为喷施外源调节物质能够快速且有效的缓解高温对水稻产生的伤害,但外源调节剂的调控效率外源生长调节剂种类、喷施浓度和喷施时间的影响较大,此方法适用于试验操作,但不适于在实际农田中进行。通过调节氮素穗肥是目前最为合理的举措之一,不仅可以有效提升颖花育性和花药蔗糖浓度,而且促进氮素向籽粒分配,进而提高产量。另外,转录组学以及代谢组学等新兴技术的崛起也为相关机理的研究提供了新思路,选育选用耐高温的水稻品种以抵御穗分化期高温,深入研究水稻中的抗逆转录因子也是研究前沿。
猜你喜欢
江西农业大学学报(2023年2期)2023-05-08
华北农学报(2022年4期)2022-09-14
作物杂志(2022年6期)2022-02-03
作物学报(2021年10期)2021-08-05
北方水稻(2021年6期)2021-02-17
东北农业大学学报(2020年12期)2021-01-15
江苏农业科学(2019年10期)2019-07-08
江苏农业科学(2016年11期)2017-03-21
广西植物(2016年10期)2016-11-11
西南农业学报(2016年4期)2016-05-17
