
多穗型与大穗型超级稻品种幼穗分化期碳氮代谢与产量构成
2022-09-14 04:22:22李文秀王礼煌方宝华王学华
华北农学报 2022年4期
李文秀,戴 力,王礼煌,方宝华,王学华
(1.湖南农业大学 农学院,湖南 长沙 410128;2.湖南省水稻研究所,农业农村部长江中下游籼稻遗传育种重点实验室,湖南 长沙 410125)
水稻是人类最重要的粮食作物,全世界以及我国均有50%~60%的人口以稻米为主食[1-2],我国是世界上最大的稻米生产国和消费国,2017年后稻米进口量开始位居世界第一[3-4]。随着经济的快速发展和城镇化水平不断提高,我国人口不断增加,耕地面积和农村劳动力不断减少、老龄化现象日益加重,单产提高对于粮食增产的重要性也不断提高。水稻的产量构成包括单位面积穗数、每穗颖花数、结实率和千粒质量[5],其中每穗颖花数是仅次于单位面积穗数(有效穗数)的重要产量贡献因子[6],也是水稻品种穗粒结构分型的重要指标因子,其多少主要取决于水稻幼穗生长发育阶段。因此,研究不同穗粒型超级稻品种的幼穗生长发育特征并针对性配套相应超高产稳产栽培技术,对于实现水稻增产、保障粮食安全具有重要意义。
大穗型品种具有单株穗数相对较少、穗粒数较多的特征,穗粒数对产量的贡献度相对较大;多穗型品种具有单株穗数相对较多而穗粒数较少的特征,单位面积穗数对产量的贡献相对较大;戴力等[7]提出的“长江中下游地区籼型超级稻穗粒分型分类系数法”将分类系数≥7的品种划分为大穗型品种、分类系数≤5的品种划分为多穗型品种。水稻有效穗数与有效分蘖数和分蘖成穗率呈正相关关系[8],穗粒数则与每穗颖花数和小花结实率正相关,而每穗颖花数又与一次枝梗、二次枝梗分化数正相关,与枝梗退化数和颖花退化数负相关[9-10];颖花退化数主要随二次枝梗的退化而发生[11]。水稻幼穗分化始期(第一苞原基分化期)是由营养生长转为生殖生长的起点,此时养分分配重点由运往分蘖开始转向运往幼穗,碳、氮代谢特点由氮代谢占优势过渡为碳、氮代谢并重,且幼穗形成过程中适宜的碳氮代谢水平有利于颖花现存数的增加,反之颖花现存数减少[10,12],不仅影响幼穗生长发育进度,还关系到产量的高低和品质的优劣[13-14]。相关研究表明,在一定范围内适当施氮有利于穗部一次枝梗、二次枝梗数的提高和颖花的形成,使二次枝梗退化数降低,二次枝梗存活率和颖花存活率增加[15-16],从而提高穗总粒数和产量[17-18]。另有研究认为,大穗型水稻穗粒数不稳定与非结构性碳水化合物NSC关系密切,且每穗颖花数与穗分化期整株碳积累量直线相关,从抽穗前16 d开始,植株体内积累的NSC越多,颖花退化率越低,如低密高氮处理可提高可溶性糖和蔗糖含量,从而促进小花的分化发育,增加可见花、可孕花数和可孕花结实率,进而提高穗粒数[19-21]。
幼穗分化期是水稻开始生殖生长的标志,更是由氮代谢为主转换为碳、氮代谢并重的关键阶段,且与穗粒数的形成密切相关。有关不同类型水稻品种各生长发育时期碳氮代谢的差异已有较多报道[22-24],但关于多穗型与大穗型品种幼穗分化阶段碳、氮代谢差异的报道不多。多穗型与大穗型品种穗粒特征形成的生物学机理不同,配套的超高产栽培技术也应不同,因此深入研究其穗粒形成差异,有利于水稻的高产稳产栽培。
本试验通过研究多穗型与大穗型品种幼穗分化阶段碳代谢生理指标(可溶性糖含量、植物淀粉酶AMS活性、蔗糖磷酸合成酶SPS活性)、氮代谢生理指标(可溶性蛋白含量、谷氨酰胺合成酶GS活性、硝酸还原酶NR活性)及产量构成的差异,探明两类品种穗粒特征形成的规律,以期为各类型品种精准配套超高产栽培技术提供理论支撑。
1 材料和方法
1.1 试验地点及材料
试验于2019,2020年在湖南省水稻研究所长沙马坡岭试验基地(28°20'N,113°09'E)进行大田小区试验。多穗型品种选用岳优9113(Y9113),大穗型品种选用天优华占(TYHZ)和五丰优T025(WT025)[7]为供试品种,其中,岳优9113根据戴力等[7]提出的“长江中下游地区籼型超级稻穗粒分型分类系数法”其分类系数为4,是典型的多穗型品种,且其适宜种植地区为广西中北部、福建中北部、江西中南部、湖南中南部、浙江南部以及湖北稻瘟病轻发区种植,范围较广;天优华占和五丰优T025的分类系数则均为8,为典型的大穗型品种。
1.2 试验设计
本试验采用随机区组设计,3个处理(品种),4次重复,共12个小区,每个小区面积为16 m2,移栽密度为20 cm×20 cm,单苗移栽。2019年于6月21日播种,7月24日移栽,2020年于6月22日播种,7月21日移栽。犁田时施用石宝山牌高浓度硫酸钾复合肥(N∶P2O5∶K2O=15∶15∶15)500.25 kg/hm2(宜昌富升化工)作基肥,移栽7 d后施分蘖肥,每公顷施用尿素(昆仑牌)75 kg和氯化钾(中化牌)35 kg。其他栽培管理措施同大田生产。
1.3 测定指标及方法
1.3.1 幼穗分化阶段的划分 分化阶段按幼穗长度法划分幼穗分化各阶段[25],具体标准见表1。
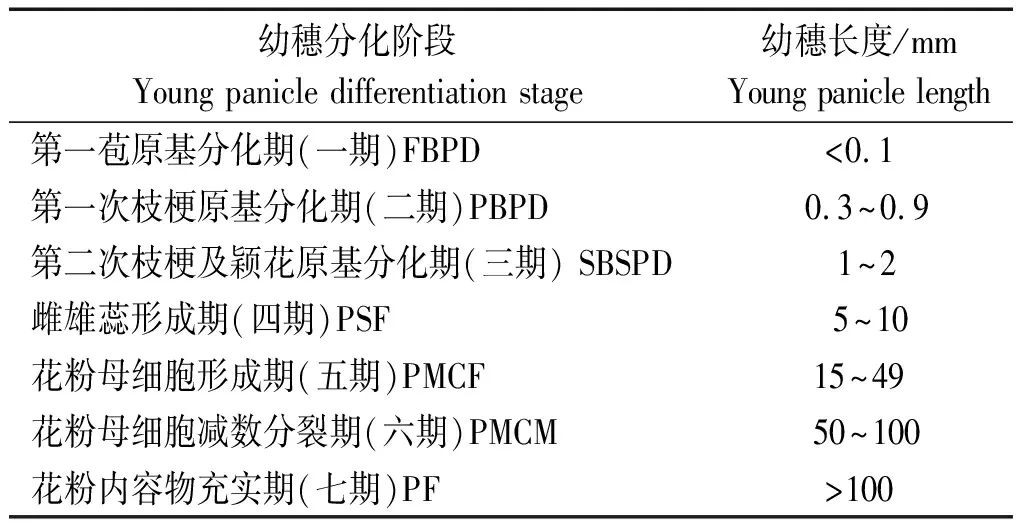
表 1 幼穗分化阶段及缩写Tab.1 Young panicle differentiation stage and acronym
1.3.2 碳代谢指标的取样方法与测定 返青后第7天开始每3 d取一次样,进行幼穗剥检观测幼穗发育情况,直至幼穗长度超过100 mm(幼穗分化七期)。每次取连续的植株主茎20根,带回室内进行幼穗剥检测定每个幼穗长度,在幼穗剥检过程中将同一分化阶段的幼穗(包括幼穗基部5 mm左右长度嫩茎)及其茎鞘和功能叶叶片(主茎最上部2片全展叶)分开保存,然后置于烘箱中105 ℃条件下杀青30 min,之后在80 ℃下烘至恒质量备用。待幼穗分化阶段完成后将同一品种不同时间取样的同一分化阶段茎鞘、叶片和幼穗样品分别汇总后粉碎备检。用植物可溶性糖检测试剂盒法(茁彩,上海茁彩生物科技有限公司)测定可溶性糖含量。
幼穗剥检确定分化阶段后,第2天再取连续植株主茎20根用冰盒带回室内进行分样。分样过程中将达到特定分化阶段的茎鞘、功能叶叶片和幼穗样品用锡箔纸分开包裹并置于液氮中,之后转移到-80 ℃超低温冰箱保存。幼穗分化阶段结束后,将同一品种不同时间取样的同一幼穗分化阶段茎鞘、功能叶叶片和幼穗分别汇总,然后制样检测。分别采用植物蔗糖磷酸合成酶(SPS)ELISA检测试剂盒法、植物淀粉酶(AMS)ELISA检测试剂盒法测定对应酶活性,试剂盒由上海茁彩生物科技有限公司提供。
1.3.3 氮代谢指标的取样方法与测定 取样方法参考碳代谢指标的方式,用植物BCA法蛋白含量检测试剂盒法(茁彩,上海茁彩生物科技有限公司)测定可溶性蛋白含量;用植物硝酸还原酶(NR)ELISA检测试剂盒法和植物谷氨酰胺合成酶(GS)ELISA检测试剂盒法测定对应酶活性,试剂盒由上海茁彩生物科技有限公司提供。
1.3.4 产量及产量构成 成熟期每个小区调查连续20穴的有效穗数并求平均值,然后根据有效穗数平均值每小区取样5穴,用于考察有效穗数、穗粒数、结实率、千粒质量和理论产量等指标;同时每小区连续割取80穴用于考察实际产量。
1.4 数据处理与分析
采用随机区组设计分析方法,用SPSS 23.0进行统计分析,Microsoft Excel 2010进行图表绘制。根据最小显著差异法(Least significant difference,LSD)检验处理间差异显著性。
2 结果与分析
2.1 多穗型与大穗型品种幼穗分化阶段不同器官的碳代谢分析
2.1.1 主茎茎鞘碳代谢分析 如图1所示,各品种幼穗分化阶段主茎茎鞘可溶性糖含量和SPS(蔗糖磷酸合成酶)、AMS(植物淀粉酶)活性均随着幼穗发育进程不断推进而增高。幼穗分化一至七期,大穗型品种(WT025和TYHZ)的主茎茎鞘可溶性糖含量高于或显著高于多穗型品种(Y9113),其中可溶性糖含量WT025分别较Y9113高31.18%(P<0.05),13.76%(P<0.05),19.45%,22.69%(P<0.05),9.52%,21.20%(P<0.05),8.97%,TYHZ分别比Y9113高18.87%,8.54%,12.65%,27.60%(P<0.05),8.30%,14.67%(P<0.05),2.69%;且大穗型品种(WT025和TYHZ)的主茎SPS活性也高于或显著高于多穗型品种(Y9113),WT025较Y9113分别高23.06%,15.86%,1.62%,20.83%,25.84%,5.21%,24.86%,TYHZ的SPS活性较Y9113分别高42.89%,19.19%,46.72%(P<0.05),8.55%,26.48%,20.78%,32.14%(P<0.05),在幼穗分化三期和七期表现显著差异(P<0.05);大穗型品种(WT025和TYHZ)的主茎AMS活性在幼穗分化一期、二期、三期和七期也分别高于或显著高于多穗型品种(Y9113),WT025分别较Y9113高7.68%,20.77%,3.92%,14.26%(P<0.05),TYHZ分别较之高8.91%,12.07%,7.73%,20.08%(P<0.05)。由此表明,大穗型品种幼穗分化阶段主茎茎鞘碳代谢水平高于或显著高于多穗型品种。
柱子上的不同小写字母表示同一幼穗分化时期不同品种处理的0.05水平上的差异。图2—6同。 Lowercase letters on the column represent significant difference at 5% probability level in the same young panicle differentiation stage with different varieties.The same as Fig.2—6.
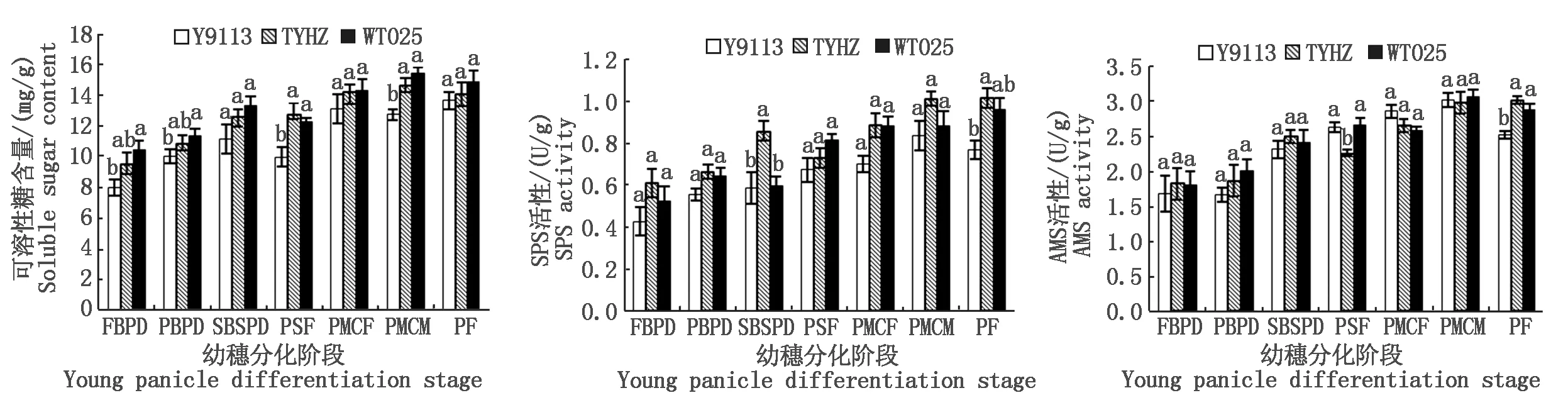
2.1.2 主茎功能叶碳代谢分析 如图2所示,幼穗分化阶段各品种主茎功能叶可溶性糖含量和碳代谢关键酶(SPS和AMS)活性均随幼穗分化推进而增高。幼穗分化一至六期,大穗型品种(WT025和TYHZ)的功能叶可溶性糖含量均高于或显著高于多穗型品种(Y9113),其中WT025分别较Y9113高13.91%,14.87%(P<0.05),8.68%,8.62%,5.44%,14.39%,TYHZ分别较Y9113高8.55%,13.68%(P<0.05),2.28%,6.96%,2.31%,7.94%;且幼穗分化一至六期,大穗型品种(WT025和TYHZ)的功能叶SPS活性高于或显著高于多穗型品种(Y9113),WT025分别较Y9113高63.02%(P<0.05),39.80%(P<0.05),28.13%,11.02%,14.96%,11.78%,TYHZ分别较Y9113高32.73%,38.19%(P<0.05),28.82%,18.00%,18.70%(P<0.05),28.55%(P<0.05);幼穗分化一至七期,大穗型品种(WT025和TYHZ)的功能叶AMS活性均高于或显著高于多穗型品种(Y9113),其中WT025一期和三期分别较Y9113高75.90%(P<0.05)和35.07%(P<0.05),TYHZ一期、三期和七期分别较Y9113高60.80%(P<0.05),23.45%,12.95%(P<0.05)。由此表明,在幼穗分化阶段大穗型品种主茎功能叶的碳代谢水平强于多穗型品种。

图2 幼穗分化阶段不同品种主茎功能叶可溶性糖含量、SPS和AMS活性Fig.2 The content of soluble sugar and the activity of sucrose phosphate synthase and amylase of main stem functional leaves at young panicle differentiation stages with different varieties
2.1.3 主茎幼穗碳代谢分析 如图3所示,幼穗分化一至七期,大穗型品种(WT025和TYHZ)的主茎幼穗可溶性糖含量高于或显著高于多穗型品种(Y9113),其中WT025分别较Y9113高16.72%,9.76%(P<0.05),35.07%(P<0.05),18.24%(P<0.05),15.56%(P<0.05),24.31%,15.22%,TYHZ分别较Y9113高3.09%,1.27%,15.21%,8.76%,21.58%(P<0.05),22.23%,14.34%;幼穗分化阶段各品种幼穗碳代谢关键酶(SPS和AMS)活性整体表现为随着生育进程的推进而逐渐增强,幼穗分化一至七期,大穗型品种TYHZ的SPS活性分别较Y9113高25.46%,18.12%,27.08%,11.55%,22.23%,19.26%,21.01%(P<0.05),且在幼穗分化一期和三期至七期,WT025的SPS活性分别较Y9113高9.36%,21.35%,5.02%,10.57%,11.43%,17.07%;在幼穗分化一至六期,各品种间的主茎幼穗AMS活性则表现为WT025分别较Y9113高7.82%,29.08%(P<0.05),4.37%,16.34%,13.44%,7.47%,TYHZ分别较Y9113高8.03%,6.99%,8.45%,1.50%,20.77%(P<0.05),10.10%。由此表明,幼穗分化阶段大穗型品种的幼穗碳代谢水平强于多穗型品种。

图3 幼穗分化阶段不同品种主茎幼穗可溶性糖含量、SPS和AMS活性Fig.3 The content of soluble sugar and the activity of sucrose phosphate synthase and amylase of main stem young panicles at young panicle differentiation stages with different varieties
2.2 多穗型与大穗型品种幼穗分化阶段不同器官的氮代谢分析
2.2.1 主茎茎鞘氮代谢分析 如图4所示,幼穗分化阶段各品种主茎茎鞘可溶性蛋白含量、NR和GS活性均随幼穗分化推进而增高,且均表现为大穗型品种(WT025和TYHZ)高于或显著高于多穗型品种(Y9113)。幼穗分化一至七期,WT025主茎茎鞘可溶性蛋白含量比Y9113分别高出13.09%(P<0.05),4.27%,7.87%,6.71%,12.83%(P<0.05),16.20%(P<0.05),13.09%(P<0.05),且在幼穗分化一期、二期和四至七期,TYHZ比Y9113分别高6.18%,6.94%,0.78%,3.15%,4.00%,8.18%;大穗型品种WT025幼穗分化四期主茎NR活性较多穗型品种Y9113显著高出27.83%,其余各阶段大穗型品种的NR活性比多穗型品种稍高或基本相当;幼穗分化一至七期大穗型品种(WT025和TYHZ)的主茎GS活性高于或显著高于多穗型品种(Y9113),其中WT025>TYHZ>Y9113,具体为WT025各时期主茎GS活性分别较Y9113高62.9%,32.03%(P<0.05),30.44%,22.50%,32.57%(P<0.05),9.43%,18.75%,TYHZ分别较Y9113高40.51%,18.08%,7.58%,21.19%,11.41%,8.91%,8.84%。由此表明,幼穗分化阶段大穗型品种主茎茎鞘氮代谢水平强于多穗型品种。
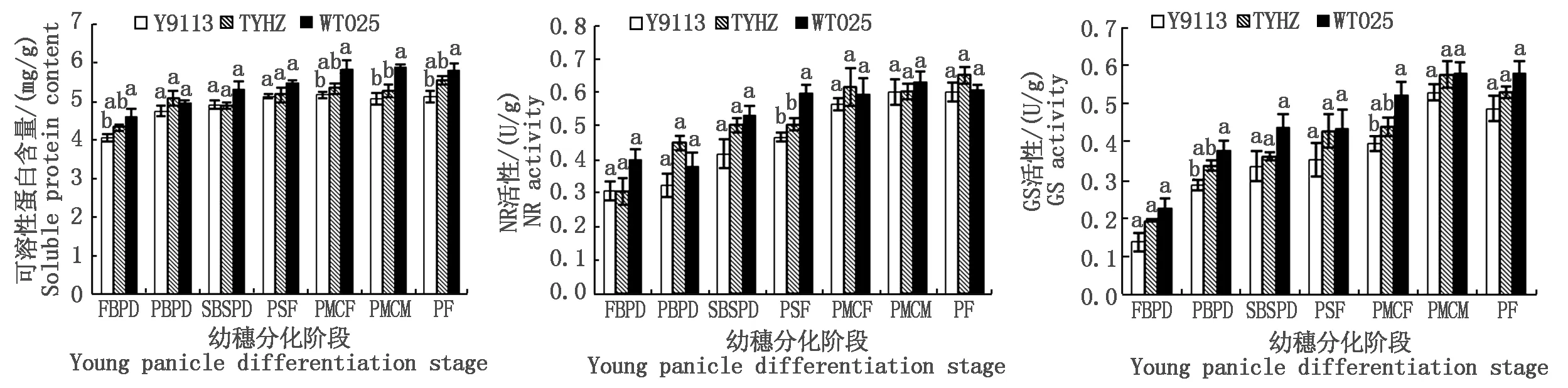
图4 幼穗分化阶段不同品种主茎茎鞘可溶性蛋白含量、NR和GS活性Fig.4 The content of soluble protein and the activity of nitrite reductase and glutamine synthesis of main stem at young panicle differentiation stages with different varieties
2.2.2 主茎功能叶氮代谢分析 如图5所示,幼穗分化阶段各品种主茎功能叶可溶性蛋白含量、NR和GS活性的变化趋势均随幼穗分化推进而增强,且其可溶性蛋白含量、NR和GS活性均表现为大穗型品种(WT025和TYHZ)高于或显著高于多穗型品种(Y9113)。具体呈现幼穗分化一至七期,WT025的可溶性蛋白含量分别较Y9113高12.87%,12.14%(P<0.05),6.77%(P<0.05),16.24%(P<0.05),22.70%,9.88%(P<0.05),28.68%(P<0.05),TYHZ分别较Y9113高9.41%,4.91%,0.80%,16.45%(P<0.05),5.26%,4.27%,11.46%(P<0.05);幼穗分化一至七期(三期除外),大穗型品种(WT025和TYHZ)功能叶NR活性高于或显著高于多穗型品种(Y9113),幼穗分化三期Y9113的NR活性稍高于WT025和TYHZ(差异不显著),其中在幼穗分化二期和五期,TYHZ的功能叶NR活性分别较Y9113高26.01%(P<0.05),30.65%(P<0.05),WT025分别较Y9113高33.40%(P<0.05),18.10%;幼穗分化一至七期,功能叶的GS活性也表现为WT025分别较Y9113高24.85%,14.92%,2.67%,63.49%(P<0.05),9.55%,10.38%,17.59%,TYHZ分别较Y9113高29.09%,4.34%,15.38%,22.04%,15.40%,16.27%,21.04%。由此说明,在幼穗分化阶段大穗型品种主茎功能叶的氮代谢水平高于多穗型品种。
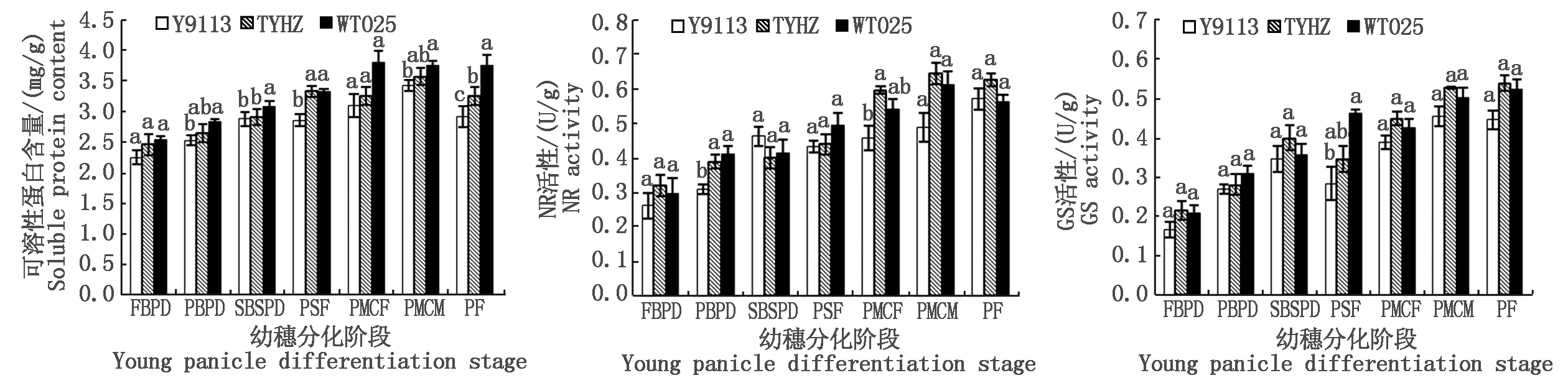
图5 幼穗分化阶段不同品种主茎功能叶可溶性蛋白含量、NR和GS活性Fig.5 The content of soluble protein and the activity of nitrite reductase and glutamine synthesis of main stem functional leaves at young panicle differentiation stages with different varieties
2.2.3 主茎幼穗氮代谢分析 如图6所示,幼穗分化一至五期,大穗型品种(WT025和TYHZ)的主茎幼穗部位可溶性蛋白含量高于或显著高于多穗型品种(Y9113),其中WT025分别较Y9113高30.02%(P<0.05),22.89%(P<0.05),11.08%,32.28%(P<0.05),13.52%,TYHZ分别较Y9113高出14.65%,13.01%,0.70%,15.60%,7.44%;且在幼穗分化一至七期,大穗型品种(WT025和TYHZ)幼穗NR活性稍高于多穗型品种(Y9113),或与之基本相当,其中一至四期和七期幼穗的NR活性WT025分别较Y9113高7.78%,15.04%,3.58%,6.31%,3.14%,TYHZ分别较Y9113高14.15%,3.68%,9.20%,4.59%,3.52%;大穗型(WT025和TYHZ)在幼穗分化一至七期,幼穗的GS活性则在某些时期表现显著差异,高于或显著高于多穗型品种(Y9113),其中WT025分别较Y9113高45.97%,50.81%(P<0.05),20.10%(P<0.05),31.24%(P<0.05),24.21%(P<0.05),16.16%,17.53%(P<0.05),TYHZ分别较Y9113高36.85%,35.98%,7.81%(P<0.05),19.69%(P<0.05),13.97%,19.68%,32.47%(P<0.05)。由此表明,幼穗分化阶段大穗型品种主茎幼穗的氮代谢水平强于多穗型品种,且在某些时期显著高于多穗型品种。

图6 幼穗分化阶段不同品种主茎幼穗可溶性蛋白含量、NR和GS活性Fig.6 The content of soluble protein and the activity of nitrite reductase and glutamine synthesis of main stem young panicles at young panicle differentiation stages with different varieties
2.3 多穗型与大穗型品种产量及产量构成差异
2.3.1 有效穗数 如表2所示,2019,2020年各品种间的单位面积有效穗数大小均表现为Y9113>TYHZ>WT025,其中,2019年Y9113较TYHZ和WT025分别高出19.2%和44.9%(P<0.05),2020年Y9113较TYHZ和WT025则分别高出29.6%(P<0.05)和52.0%(P<0.01)。
2.3.2 穗粒数 2019,2020年各品种间的穗粒数大小则均呈WT025>TYHZ>Y9113的现象,2019年WT025和TYHZ分别比Y9113极显著高出60.9%(P<0.01)和54.0%(P<0.01),2020年则分别比Y9113极显著高出67.3%(P<0.01)和42.7%(P<0.01)。
2.3.3 结实率 从表2可以看出,2019年各品种的结实率大小表现为WT025>Y9113>TYHZ,2020年9月中下旬和10月上中旬由于遇上“寒露风”连续低温影响各品种抽穗灌浆,导致各品种的结实率均出现下降现象,2020年岳优9113的结实率与2019年相比下降了21.43百分点,TYHZ的结实率与2020年相比下降了7.96百分点,WT025的结实率与2020年相比下降了7.27百分点。由此可明显看出,在遇“寒露风”天气时,多穗型Y9113的结实率下降程度大于大穗型品种TYHZ和WT025。
2.3.4 千粒质量 2019,2020年各品种千粒质量大小均表现为Y9113>TYHZ>WT025,且各品种间均表现出极显著差异,2019年Y9113 较TYHZ和WT025分别极显著高出4.4%(P<0.01)和15.2%(P<0.01),2020年Y9113较TYHZ和WT025分别极显著高出7.1%(P<0.01),15.6%(P<0.01)。
2.3.5 产量 2019年各品种间的理论产量未表现出显著差异,2020年的理论产量则表现出WT025高于TYHZ,并显著高于Y9113,较Y9113高出21.9%(P<0.05);各品种间的实际产量大小2019,2020年均呈现WT025>Y9113>TYHZ的规律,且在2020年WT025的实际产量较Y9113极显著高出20.2%(P<0.01),TYHZ与Y9113之间无显著差异。
上述结果表明,多穗型品种的主要特征表现为单位面积有效穗数相对较多,穗粒数相对较少的特征,大穗型品种则主要表现为单位面积有效穗数相对较少,穗粒数相对较多的特征。
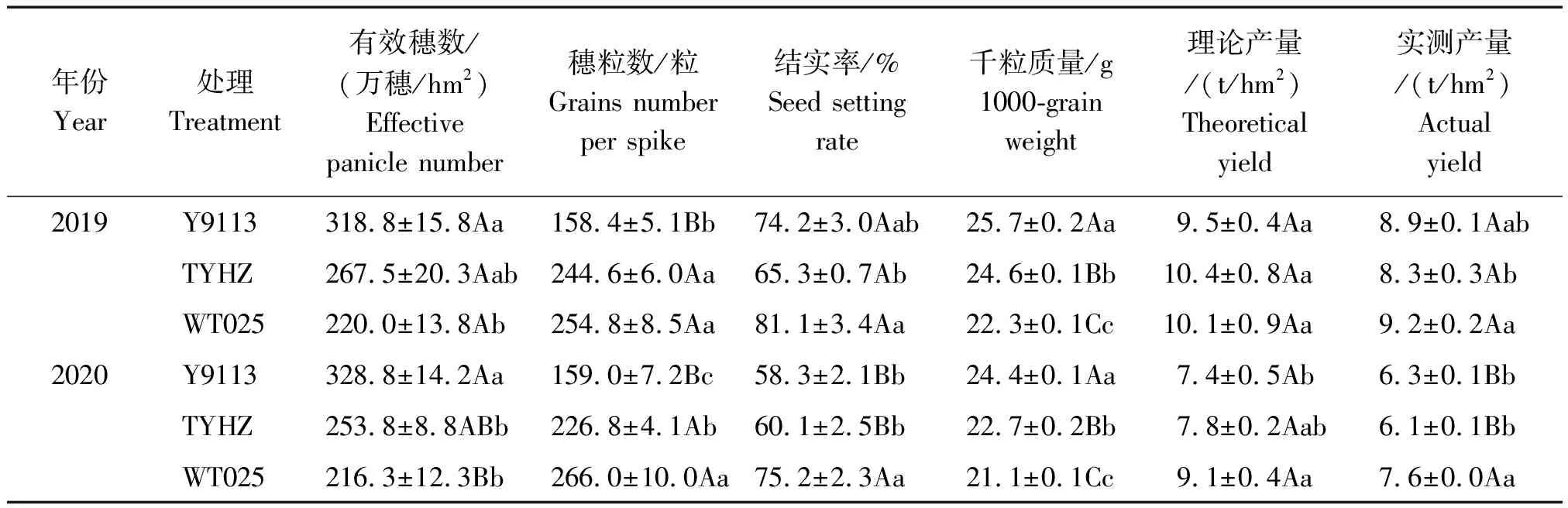
表2 多穗型与大穗型品种产量及产量构成因素Tab.2 Yield and yield components of multi-panicle type and large-panicle type varieties
2.4 幼穗分化期碳氮代谢与产量性状相关性分析
幼穗分化时期各器官可溶性糖和可溶性蛋白含量与产量性状指标间相关分析结果显示,穗粒数与幼穗分化二期主茎茎鞘(r=0.935,P<0.01)和功能叶(r=0.866,P<0.05)可溶性糖含量及四期幼穗可溶性蛋白含量(r=0.867,P<0.05)呈显著或极显著正相关;有效穗数与二期茎鞘可溶性糖呈显著负相关(r=-0.818,P<0.05);千粒质量与三期茎鞘可溶性糖含量呈显著负相关(r=-0.909,P<0.05);实际产量则与幼穗二期至七期的可溶性蛋白含量均呈显著负相关(P<0.05),其相关系数分别为-0.906,-0.842,-0.874,-0.841,-0.819,-0.843。表明幼穗分化二期(第一次枝梗原基分化期)和三期(第二次枝梗原基和小穗原基分化期)的可溶性糖和可溶性蛋白代谢水平对水稻穗粒结构的形成具有重要意义。相关代谢酶活性与产量性状指标的相关性有待进一步研究。
3 结论与讨论
3.1 多穗型与大穗型品种幼穗分化阶段碳代谢分析
碳代谢在整个植物生理代谢中都有着非常重要的地位,碳水化合物是植物体内多种生长发育和生理生化反应的产物,也是能量的来源,是重要的研究指标[26],植物的碳代谢过程主要包括光合碳同化、蔗糖和淀粉的合成与代谢、碳水化合物的运输和利用,与植物碳代谢密切相关的酶主要有蔗糖合成酶(SS)、蔗糖磷酸合成酶、转化酶(Inv)以及淀粉酶等,SPS调节光合产物在蔗糖和淀粉的分配,是植物体内蔗糖合成、碳同化与分配的一个关键调节酶[27-28],且SPS活性与可溶性糖含量呈正相关[29]。本研究结果发现,水稻幼穗分化各阶段主茎茎鞘、功能叶和幼穗的SPS活性、AMS活性和可溶性糖含量大体均表现为大穗型品种高于或显著高于多穗型品种,表明大穗型在幼穗分化阶段的碳代谢水平高于多穗型品种,且在第二次枝梗和颖花原基分化期(幼穗分化三期)和花粉母细胞减数分裂期(幼穗分化六期)分别是粒数快速增加以及枝梗和颖花易发生死亡或退化的时期,与大穗的形成紧密相关[30-32],在本研究中,发现在第二次枝梗和颖花原基分化期,大穗型品种天优华占主茎的SPS活性较岳优9113显著高出46.72%,大穗型品种五丰优T025幼穗的可溶性糖含量较岳优9113显著高出35.07%,以及在花粉母细胞减数分裂期,大穗型品种五丰优T025主茎可溶性糖含量较岳优9113显著高出21.20%,大穗型品种天优华占的主茎茎鞘可溶性糖含量和功能叶SPS活性较岳优9113分别显著高出14.67%和28.55%。且根据幼穗分化期碳氮代谢与产量性状相关性分析可知,穗粒数与幼穗分化二期主茎茎鞘和功能叶可溶性糖含量呈显著或极显著正相关,有效穗数与二期茎鞘可溶性糖含量呈显著负相关。前人研究表明,叶鞘和茎秆积累的淀粉和可溶性糖含量增多,输送给幼穗构建花器官的碳水化合物也多,能促进枝梗和颖花的分化变退化为不退化或少退化[33]。由此可见,大穗型品种穗粒数多的形成机制与旺盛的碳代谢有关,大穗型品种较高的SPS活性、AMS活性和可溶性糖含量导致大穗型品种能合成更多的碳水化合物供给枝梗和颖花的分化,为保证大穗型品种的穗粒数提供了基础。
3.2 多穗型与大穗型品种幼穗分化阶段氮代谢分析
氮代谢包括氮素在植物体内的吸收、同化、转运、利用和调节等过程,NR和 GS是影响植物氮素代谢的关键酶,其活性强弱直接影响着氮素同化速率的高低,反映了植株营养状况和氮素代谢水平[26,34-35]。有研究指出,不同穗型品种间植株含N率差异较小,但大穗型品种抽穗期群体、个体氮素累积量均较大[36],在幼穗期通过追施氮素营养能增加二次枝梗上分化的颖花数和减少二次枝梗上的退化颖花数,提高每穗颖花分化数,在枝梗和颖花分化期供应充足有利于大穗的形成,在雌雄蕊分化期氮素营养可以不同程度的增加小花分化数,而在减数分裂期可以显著减少颖花退化数[37]。另有研究表明,各生育期水稻叶片NR、GS、GOGAT 活性随着氮含量的增加而提高[38],NR 和 GS 活性与蛋白质含量呈正相关[39-41]。在本研究中,大穗型品种幼穗分化阶段主茎茎鞘、功能叶和幼穗的NR和GS的活性大体高于或显著高于多穗型品种,其可溶性蛋白质含量也表现为大穗型品种高于或显著高于多穗型品种,且在第二次枝梗和颖花原基分化期,大穗型品种五丰优T025功能叶的可溶性蛋白质含量和幼穗的GS活性较多穗型品种岳优9113分别显著高出6.77%和20.10%,大穗型品种天优华占幼穗的GS活性较岳优9113显著高出7.81%,以及在花粉母细胞减数分裂期,大穗型品种五丰优主茎茎鞘和功能叶的可溶性蛋白质含量较岳优9113分别显著高出16.20%和9.88%。根据幼穗分化期碳氮代谢与产量性状相关性分析可知,穗粒数与四期幼穗可溶性蛋白含量呈显著正相关。由此可见,大穗型品种在幼穗分化阶段其主茎茎鞘、功能叶和幼穗的氮代谢均强于多穗型品种,且在幼穗分化阶段大穗型品种较强的氮代谢使大穗型品种在幼穗分化阶段能获得更多的枝梗和颖花分化数,从而促进大穗型品种形成较多的穗粒数。
大穗型水稻品种在幼穗分化一至七期的碳代谢与氮代谢均强于多穗型品种,有利于枝梗和颖花的分化以及减少颖花退化,促进穗粒数的增加,是大穗型品种形成大穗的生理机制之一,也是多穗型品种与大穗型品种穗粒形成特征的差异之一。
猜你喜欢
Acta Mathematica Scientia(English Series)(2024年1期)2024-03-23 08:02:54
江西农业大学学报(2023年2期)2023-05-08 01:32:10
作物杂志(2022年6期)2022-02-03 04:56:22
热带作物学报(2021年9期)2021-11-08 11:55:35
作物学报(2021年10期)2021-08-05 11:30:40
北方水稻(2021年2期)2021-04-26 08:25:56
核农学报(2021年3期)2021-02-22 08:47:58
北方水稻(2021年6期)2021-02-17 07:17:50
茶叶科学(2018年5期)2018-10-17 01:56:42
湖北农业科学(2017年6期)2017-04-26 14:37:05
