
整合素通过PI3K/Akt 信号通路调控畜禽骨骼健康的研究进展
2024-03-21 06:04路亚男丁文丽岳涛静徐博文黄淑成
畜牧与饲料科学 2024年1期
路亚男,丁文丽,岳涛静,徐博文,黄淑成
(河南农业大学动物医学院,河南 郑州 450046)
随着近年来国内外畜禽业的迅速发展, 由于生产养殖的集中化以及饲养管理和基因遗传控制的改善,畜禽的生长周期大大缩短,导致畜禽体重在短期内迅速增加,骨骼的强度不足以支撑机体的快速发展,造成畜禽骨骼长期处于亚健康状态[1]。骨骼是脊椎动物中坚硬的结缔组织, 具有构成机体基本支架、保护脏器、支撑体重和维持运动的作用[2]。 骨骼发育主要是通过膜内成骨和软骨内骨化完成的, 该过程由多种调控因子组成的复杂调控网络精准调控破骨细胞和成骨细胞生成以确保骨代谢正常。 成骨细胞(osteoblast)是骨形成细胞的重塑单位,是间充质干细胞的终末分化产物,成熟的成骨细胞是位于骨表面的单层细胞, 具有合成骨基质的重要功能[3]。破骨细胞(osteoclast)主要分布在骨表面,通过释放乳酸、柠檬酸、碳酸、碳酸酐酶以及溶酶体等对骨组织进行分解破坏。 在骨重建过程中,破骨细胞贴附在旧骨区域,分泌酸性物质溶解矿物质,分泌蛋白酶消化骨基质,形成骨吸收陷窝;随后,成骨细胞移行至被吸收部位,分泌骨基质,骨基质矿化后形成新骨[4]。
骨的分解吸收与新骨形成之间的平衡是维持正常骨代谢的关键。 骨代谢是指机体内骨骼组织在生长、发育和维持骨量平衡过程中,成骨细胞与破骨细胞进行的一系列维持机体正常骨量的过程。 其中,成骨细胞负责合成和矿化新骨,破骨细胞负责对矿化的骨基质进行分解吸收, 生长时期骨量呈线性增长,骨生长停止后,破骨细胞对骨的吸收和成骨细胞的生成协同作用使骨骼处于平衡状态[4]。 骨代谢通常发生在骨表面的聚焦部位,即骨代谢单位(bone metabolic unit,BMU),许多BMU的累积行为构成整个机体的骨代谢, 如果周转率增加, 骨形成组织中的缺陷或骨吸收的任何不平衡都有可能导致功能完整性发生明显变化。 骨形成受损会导致骨质疏松中的骨量降低和骨骼脆性升高, 临床上骨代谢疾病主要包括佝偻病、 软骨病、骨质疏松症等,主要表现为骨折、骨痛、肌无力和发育畸形[5]。
近几年,人们对于整合素(integrin)通过磷脂酰肌醇3-激酶/蛋白激酶B(phosphatidylinositol 3-kinase/protein kinase B,PI3K/Akt)信号通路影响骨骼健康的关注度增加。越来越多的研究表明,整合素β1 和β3 参与成骨细胞分化过程中各种细胞因子调节过程[6]。 整合素作为PI3K 的上游引物之一, 其中几种亚基在受到胞外物质的刺激后自身发生磷酸化,而后通过PI3K/Akt 途径引起骨代谢发生变化。 有研究指出, 整合素家族中的整合素α2β1 被证实通过细胞外信号调节激酶(extracellular signal-regulated kinase,ERK) 途径在促进成骨细胞分化中发挥重要作用[7]。 整合素αvβ3 与集落刺激因子1 受体 (colony stimulating factor 1 receptor,CSF1R 或c-FMS)结合,形成破骨细胞迁移所需的细胞骨架, 通过激活ERK/c-Fos 信号通路来调节细胞黏附、分化和增殖[8-9]。 研究表明,富血小板血浆可以通过激活整合素改变局部黏着斑激酶 (focal adhesion kinase,FAK) 的结构, 再通过PI3K/Akt 信号通路治疗兔牙槽骨骨内缺损, 改善骨愈合的过程[10]。 整合素可能通过PI3K/Akt 信号通路对畜禽骨代谢发挥调控作用, 但具体机制仍需要进一步研究。 整合素、PI3K/Akt 信号通路、畜禽骨健康三者之间存在紧密的联系, 通过干预整合素-PI3K/Akt 信号通路来维持骨骼健康具有潜在的开发前景。综合国内外文献报道,笔者在阐述整合素和PI3K/Akt 信号通路对骨代谢作用的基础上,重点综述了整合素通过PI3K/Akt 信号通路影响畜禽骨代谢的研究进展, 以期为预防和治疗畜禽骨骼疾病提供参考。
1 整合素
1.1 整合素概述
整合素是由α 亚基和β 亚基以非共价键结合形成的异源二聚体,在细胞膜表面参与细胞之间、细胞与胞外基质之间相互作用的一种跨膜细胞黏附分子。 在哺乳动物中, 目前发现整合素至少有18 种α 亚基和8 种β 亚基, 组成24 种整合素分布在机体不同的组织器官中[11]。 整合素的2 条支链在空间上平行于细胞外端汇合形成“U”字形结构,结构上包括大的细胞外区、单一的单次跨膜区和很短的胞内区三部分[12]。
整合素的功能主要是通过其在细胞膜表面发生的非活化构象-活化构象之间的变化以及胞内转运等多种机制共同调节发挥作用。 具体表现为整合素的细胞质结构域协调细胞骨架聚合物和细胞内复合物的组装, 细胞外结构域与相邻细胞表面的受体或细胞外基质大分子结合[13],从而介导细胞生存、免疫监视、血管生成、发育和炎症等多种生理过程。整合素功能异常会引起免疫缺陷、炎症性疾病、骨代谢异常、肿瘤和癌症等疾病[14]。 此外,整合素可激活FAK/PI3K/Akt 通路,即作为刺激因子通过FAK/PI3K/Akt 信号通路对骨骼健康造成一定程度的影响。
1.2 整合素活性调控
已有研究表明,激素、细胞因子、细胞外基质(extracellular matrix,ECM)等刺激因子可以激活整合素,使其蛋白发生磷酸化,导致细胞内Ca2+水平和pH 值升高,产生磷脂代谢产物以进行后续的信息传递[14]。 整合素可以将细胞外信号向细胞内传导,也可以将细胞内信号向细胞外传导,这是其独有的双向信号传导的特性。 整合素信号的传递恰好体现了其活化与非活化状态的转变, 这与整合素构象直接相关, 而上调蛋白和配体结合的过程就是整合素的活化过程[15]。在正常生理状态下,整合素以非活化的折叠构象存在于细胞表面,α、β亚基结构域紧密排列, 胞内结构域弯曲且彼此靠近,处于亲和力较低的状态。当整合素被刺激因子激活后,尾部结构域完全直立,α、β 亚基头部结构域完全伸展;尾部结构域进一步分离,hybrid 结构域外摆并改变尾部构象,头部结构域打开,胞内结构域不再是紧密联系而是彼此分开。一方面,胞内调控蛋白与整合素α、β 亚基胞内段结合, 通过由内向外的信号使整合素转变为活化构象; 另一方面, 胞外结构域与细胞外基质配体以高亲和力结合, 引发整合素构象进一步伸展及其在细胞膜上的聚簇, 通过由外向内的信号激活胞内信号通路(见图1)[16]。
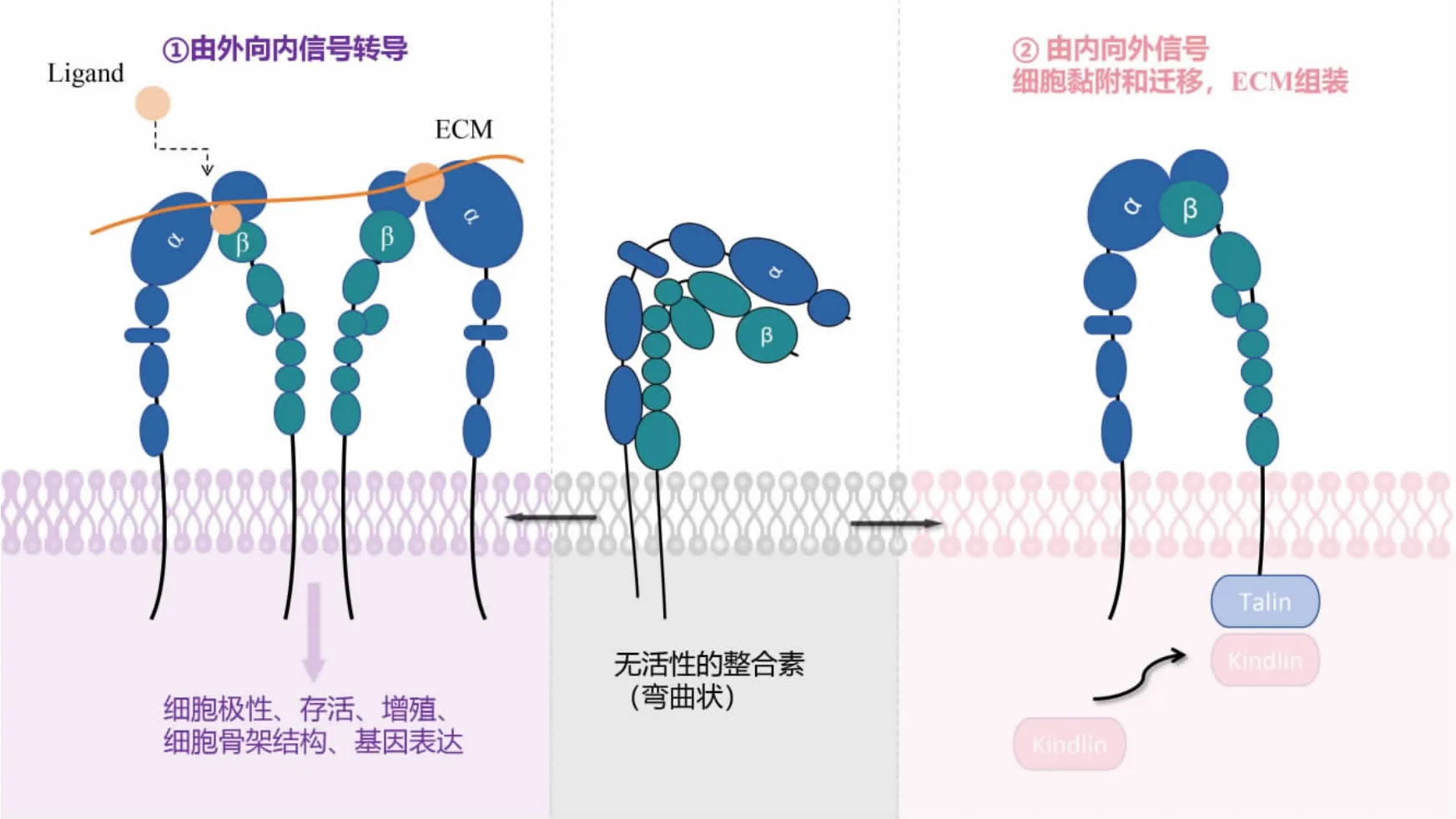
图1 整合素活化机制
在由外向内的信号转导过程中,配体(Ligand)与整合素受体蛋白结合,最后结合在整合素上;踝蛋白(Talin)是一种与细胞迁移运动高度相关的蛋白质,常见于FAK 结构内;黏着斑蛋白Kindlin 是一种新型的、在进化上高度保守的、含有FERM 结构域的蛋白家族。
1.3 整合素在骨骼健康中的作用
整合素广泛表达于全身各组织器官, 参与机体内多种生理功能的调控过程。 整合素及其活性调控相关蛋白和因子的表达缺失会影响组织的形成,造成不同类型疾病的发生。在整合素与骨骼健康的相互关系中, 最常见的是整合素参与调节骨关节炎的病理过程, 具体表现主要是整合素分布在软骨中,某些亚基可以与层粘连蛋白结合,从而在骨关节炎中发挥重要作用[17]。 整合素α5β1 的α5 亚基缺失可影响软骨损伤,α5β1 的亚基传导过量的应激刺激引起细胞外基质损伤与骨关节炎的发生有关[18]。整合素αvβ3 可促进肿瘤细胞侵袭和骨细胞溶解[19]。 整合素活化过程中由于抑制蛋白作用受阻而引起的疾病不多, 有报道表明,filamin 基因突变主要引起骨骼病变,说明filamin 蛋白抑制后导致整合素无法活化, 这对于骨代谢也产生极大影响[20-21]。 此外,有研究表明整合素也与癌症、炎症有关。 整合素β4 在肝癌细胞中高度表达, 运用信号通路抑制剂PKI-587 能够逆转肝细胞癌增殖、迁移的效应[22]。 Kindlin-3 缺失使整合素迁移能力下降,进一步影响细胞黏附,可能导致机体出现炎症和免疫反应等[23]。 目前国内外对于整合素在骨骼健康方面的研究还主要集中在骨关节炎上, 关于整合素对其他骨骼疾病的调控作用机制有待进一步研究。
2 PI3K/Akt 信号通路对骨骼健康的调控机制
PI3K/Akt 信号通路是由PI3K 和Akt 构成的重要通路。 PI3K 是生长因子受体蛋白酪氨酸激酶2(protein tyrosine kinase 2,PTK2)下游的主要信号传导成分, 属于脂质酶家族, 其活化由质膜上PTK2 的磷酸化触发[24]。磷酸化脂质信号的传导发生在细胞膜上, 有助于各种信号传导成分的募集和激活。 Akt 又称为丝氨酸/苏氨酸蛋白激酶B(PKB),作为PI3K 最重要的下游效应器,在整个级联信号转导中发挥重要作用。 哺乳动物中Akt分 为3 种 亚 型:Akt1/pkBα、Akt2/pkBβ 和Akt3/pkBγ, 因其亚型特异性可被多种信号激活[25-26]。Akt 通 过3,4,5- 三 磷 酸 磷 脂 酰 肌 醇(phosphatidylinositol 3,4,5-trisphosphate,PIP3) 与Akt PH 区结合而被激活,引起构象发生改变,进一步导致Ser473 和Thr308 位点双磷酸化,Akt 从细胞膜转移至其他细胞区域进行磷酸化, 进一步激活下游效应分子。 PI3K-Akt 信号通路调节许多正常细胞过程,主要在细胞增殖、存活、生长、运动和炎症反应等多个过程发挥关键作用[27]。
有研究报道, 利用玫瑰树碱及自体骨髓间充质干细胞对软骨降解模型兔进行治疗, 发现炎症因子减少、 基质金属蛋白酶 (matrix metalloproteinase,MMP)下调,从而实现对软骨的保护作用,此外,PI3K/Akt、p38 丝裂原活化蛋白激酶(p38 mitogen-activated protein kinase,p38 MAPK)、核因子κB(nuclear factor kappa-B,NF-κB)等炎症通路下调可减轻炎症[28];推测玫瑰树碱和自体骨髓间充质干细胞治疗通过PI3K/Akt 信号通路减轻炎症,进而可能通过下调MMP 来促进退化关节软骨的修复。 在骨肿瘤的治疗中需要面对癌细胞残留和术后骨缺损的巨大挑战,而氨基功能化锆基MOF纳米颗粒有可能通过PI3K/Akt 信号通路或者MAPK 途径增强前成骨细胞的成骨分化作用[29]。据报道,PI3K/Akt 通路与成骨细胞和骨髓间充质干细胞的凋亡密切相关。 杯苋甾酮可能通过骨髓间充质干细胞中的PI3K/Akt 信号通路抑制地塞米松诱导的细胞凋亡的激活, 可能通过减少空骨腔隙的百分比促进类固醇诱导的股骨头坏死(steroid-induced osteonecrosis of the femoral head,SIONFH)模型大鼠的骨修复[30]。
PI3K/Akt 信号通路是与骨骼健康密切相关的经典信号通路,通过炎症、自噬、凋亡等作用调节骨代谢平衡。 利用3,4-二羟基苯丙氨酸-IGF-1(3,4 -hydroxyphenalyalanine -containing recombinant insulin-like growth-factor-1,DOPA-IGF-1)修饰包裹小檗碱的聚乳酸-乙醇酸-羟基磷灰石(poly lactic-co-glycolic acid/hydroxyapatite,PLGA/HA)微球,封装在复合微球中的小檗碱可以用于骨缺损的临床治疗,增强骨缺损修复效果,同时,体外试验结果表明小檗碱可以有效激活IGF-1R/PI3K/Akt/mTOR 通路增强胰岛素样生长因子-1(insulin-like growth factor-1,IGF-1)介导的成骨分化作用[31]。 左归丸能通过PI3K/Akt 通路提高血液中骨桥蛋白的含量, 调节骨保护素(osteoprotegerin,OPG)/核因子κB 配体的受体激活物(receptor activator of nuclear factor kappa-B ligand,RANKL)(OPG/RANKL)等多种通路,增加骨密度并改善 骨 微 结 构[32]。 黄 酮 类 化 合 物Boeravinone B(BOB)可以完全抑制RANKL 诱导的PI3K/Akt 信号级联反应,说明BOB 可以通过抑制PI3K/Akt 途径来改善骨量, 这对于破骨细胞和脂肪细胞的分化至关重要。 另外,BOB 还可以通过调节NF-κB和MAPK 通路抑制破骨细胞分化[33]。 通过以上研究可知,PI3K/Akt 信号级联反应可以通过不同的上下游信号对骨骼发挥作用, 最终引起骨代谢发生一系列变化。 然而,目前关于PI3K/Akt 信号通路在畜禽骨骼调控方面的研究还较少。
3 整合素通过PI3K/Akt 信号通路对畜禽骨代谢的作用机制
整合素可以接收来自细胞内和细胞外的信号,被激活时自身发生磷酸化,FAK 通过整合素受体跨膜连接到ECM,在质膜的细胞质内侧形成黏斑[34]。整合素需要对ECM 配体表现高亲和力的开放构象, 在FAK 及其复合物磷酸化的状态下,才可实现高效的细胞信号迁移。详细过程如下:胞外配体与整联蛋白结合后,FAK 分子中的酪氨酸激酶Src 活化, 磷酸化的FAK 暴露Tyr-397 位点,PI3K 的SH2 结构域直接与Src 形成复合体,活化PI3K,活化的磷脂酰肌醇产生2 种衍生物,即PI-3,4-二磷酸和PI-3,4,4,5-三磷酸[34]。
PI3K 和Akt 是FAK 的下游基因,在磷酸化状态下被FAK 激活,FAK 又受上游调控因子整合素的调控。 Casar 等[35]研究发现活性整合素β1 与激活状态下的CUB 结构域蛋白1(CUB domain containing protein 1,CDCP1)复合,CDCP1 的整联蛋白β1 与CDCP1 片段复合诱导细胞内磷酸化信号传导。 Luo 等[36]研究发现乳腺癌患者血清中的14,15- 环 氧 二 十 碳 三 烯 酸 (14,15-epoxyeicosatrienoic acid,14,15-EET)可上调整合素αvβ3的表达水平,导致FAK/PI3K/Akt 信号激活,通过整合素αvβ3 和FAK/PI3K/Akt 级联反应激活诱导乳腺癌细胞对EMT 和顺铂耐药。 Oudart 等[37]证明胶原蛋白XIX 的NCI 结构域通过诱导整合素αvβ3 磷酸化来控制FAK/PI3K/Akt/mTOR 信号途径,这对于黑色素瘤的治疗具有积极影响。
畜禽骨骼的生长发育受多种微环境因素的调节,包括整合素和细胞外基质复合体的相互作用。参与骨骼发育过程的主要细胞包括骨细胞、 成骨细胞、破骨细胞、软骨中的软骨细胞,骨代谢发生异常时, 有可能是受到生物力学损伤或应激源刺激, 这些因素无论是单独作用还是组合作用都可能与骨骼健康程度有密切关系[38]。有报道表明,对患骨关节炎的马用整合素α10β1 选择性间充质干细胞治疗后, 其跛行和腕关节弯曲症状随着治疗时间的推移有所好转, 说明该疗法可以减轻骨关节炎和软骨退化[39]。 骨关节炎患者关节滑液中整合素β2 的高水平表达可作为辅助诊断骨关节炎的指标[40]。 细胞外基质中的层粘连蛋白通过与整合素结合相互作用导致成骨能力降低,在DBM 支架添加非胶原性层粘连蛋白CBD-LN 肽能够明显促进骨髓间充质干细胞和内皮祖细胞的增殖[41]。以上研究体现了整合素对于畜禽骨骼肌在不同方面的作用, 然而整合素发挥作用是否经过下游机制PI3K/Akt 通路则需要进一步探讨。
影响骨骼健康的信号通路有很多, 如骨形态发生蛋白(bone morphogenetic protein,BMP)/果蝇母本抗生存因子蛋白 (small mother against decapentaplegic protein,Smad)(BMP/Smad) 和p38 MAPK 通路,以及PI3K/Akt、Wnt/β-连环蛋白(βcatenin)、ERK、应激激活蛋白激酶(stress-activated protein kinase,SAPK)、NF-κB 诱导激酶通路[42]。笔者重点关注的是PI3K/Akt 通路影响畜禽骨骼健康的相关机制。 整合素通过PI3K/Akt 信号途径引起的机体相应变化与细胞增殖、迁移、肿瘤、癌症和骨骼代谢都有密切联系。有报道指出,肠道微生物菌落失调也与代谢性骨病有关。 高血糖可抑制PI3K/Akt 通路,诱导血管收缩,导致血管内皮细胞损伤,而血管的损伤可能会导致成骨细胞、破骨细胞和间充质干细胞营养不足, 最终导致骨发育不良[1]。 强直性脊柱炎患者机体伴随骨侵蚀引起的骨质疏松,青藤碱可以通过细胞膜上的整合素α5调控下游PI3K/Akt 信号通路来调控成骨分化,而过表达整合素α5 使成骨分化上调,敲低则会抑制成骨分化[42-43]。 由此可知, 临床上可通过调控PI3K/Akt 信号通路影响成骨细胞的分化, 这可能为将来治疗畜禽骨骼疾病提供新思路。
3.1 整合素通过PI3K/Akt 信号通路对畜禽骨形成过程的影响
整合素参与畜禽骨代谢的多个环节, 对于畜禽骨代谢具有双向调节作用,不同亚基、不同类型的整合素参与特定的生理、病理过程,作为重要的信号转导分子, 通过介导多种生物物理刺激来调节骨代谢。Liu 等[43]利用兔下颌骨人工缺损模型研究发现,富血小板血浆可通过激活整合素,使其改变FAK 结构,进一步触发PI3K/Akt 通路来提高骨再生和骨沉积的速度。然而PI3K 抑制可导致充间质祖细胞的增殖和成骨分化水平显著降低, 从而减少骨形成。 Wang 等[44]证明成骨细胞中BK 蛋白(尤其是BKα 的C 端)与整合素β1 之间存在相互作用,可通过FAK 和ERK 激活整合素信号传导,随后导致转录因子Runx2 表达水平增加, 加速成骨细胞分化和骨形成。 Qin 等[45]研究表明,小鼠骨细胞中整合素β1 表达的缺失会导致承重长骨和脊柱的骨量严重降低,以及骨细胞腔隙-小管系统增大和细胞形态异常。总而言之,整合素磷酸化结构被激活时, 可以使骨形成增加。 但也有报道称ATN-161 能对整合素α5β1 起到直接的抑制作用,减少MMP-13 的表达,减轻软骨的降解,下调B 细胞淋巴瘤-2 相关X(Bax)蛋白,上调B 细胞淋巴瘤-2(B-cell lymphoma 2,Bcl-2)蛋白,从而抑制软骨细胞凋亡,进一步保护关节软骨,减缓关节软骨的退变[46]。
FAK 作为PI3K 上游重要的信号分子之一,被质膜上的整合素和生长因子受体激活后将信号分子通过PI3K/Akt 信号通路传递至靶点,其在骨骼健康中发挥着十分重要的作用, 能够在谱系分化过程中协调细胞骨架的变化。 He 等[47]研究表明,BMP9 可作为成骨诱导剂诱导骨髓间充质干细胞成骨分化,促进骨缺损修复,还可通过BMP/Smad、环氧化酶-2 (cyclooxygenase-2,COX2)、 视网酸、IGF2 和FAK 来调节多种骨髓间充质干细胞的成骨分化; 分泌的酸性和富含半胱氨酸的蛋白质(secreted protein acidic and cysteine rich,SPARC)相关模块钙结合蛋白2 (SPARC related modular calcium binding protein2,SMOC2) 可 通 过FAK/PI3K/Akt 途径来增强BMP9 诱导的成骨分化,这可能通过促进SMOC2 和整合素β1 的相互作用来触发。 Schreiber 等[48]研究证实小鼠二磷酸腺苷核糖化因子鸟苷酸激酶1 (adenosine diphosphate ribosylation factor guanylate kinase 1,ASAP1)缺失能通过FAK/Src 和PI3K/Akt 信号传导引起生长迟缓和骨化延迟, 具体表现为生长板肥大区扩大和软骨骨连接紊乱, 同时还会导致体外间充质细胞的软骨形成加速,抑制骨和脂肪形成分化。以上研究证实,PI3K/Akt 是传导调节骨形成作用的重要信号通路。
进一步来讲, 整合素受到细胞内外物质的刺激时,可能通过PI3K/Akt 传导途径引起骨代谢发生一系列反应。 Lee 等[49]研究发现,流体剪切力能够刺激骨细胞生成和释放成骨细胞因子, 通过骨细胞互联网络的间隙连接转移, 进而诱导成骨细胞募集和骨细胞生长。 振荡剪切应力(oscillatory shear stress,OSS)诱导的成骨样细胞MG63 增殖是由整合素αvβ3 和β1 通过与FAK 和Shc 介导发挥协同作用形成与PI3K 的异聚复合物,以及响应OSS 刺激引起下游的PI3K-Akt/ERK 引发信号级联。 实际上,在响应OSS 的MG63 细胞中,PI3K 是激活Akt 和ERK 所必需的。 也有研究表明,对小鼠给予治疗性低强度脉冲超声后, 成骨细胞和骨细胞标志蛋白Wnt1 和β-catenin、 破骨细胞标志蛋白RANKL 以及抗凋亡蛋白Bcl-2 的表达显著上调; 下颌成骨细胞受到机械负载后经过整合素α5β1/PI3K/Akt 通路促进其重塑并提高成骨细胞存活能力。在缺乏低强度脉冲超声的情况下,颅骨Bcl-2、 整合素连接激酶 (integrin-linked kinase,ILK) 以及磷酸化的哺乳动物雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR) 均完全表达[50]。 综合以上研究表明,整合素通过PI3K/Akt来调控畜禽骨的形成(见图2)。

图2 整合素通路PI3K/Akt 信号通路对骨骼健康的调控机制
3.2 整合素通过PI3K/Akt 信号通路对畜禽骨吸收过程的影响
整合素在信号传导过程中受到某些刺激因子的作用时也可以对畜禽骨吸收方面产生一定的影响。骨吸收是指在较低的应力水平下,骨组织的体积和密度逐渐发生下降的生理行为。 力学行为与破骨细胞明显相关,当应力水平较低时,破骨细胞比成骨细胞活跃则会引起骨的总量降低。Qin 等[51]研究表明骨细胞整合素β3 的表达缺失会导致成年小鼠的承重长骨骨量减少, 骨细胞中树突的数量和长度减少。骨细胞整合素β3 的表达缺失显著降低了成骨细胞介导的骨形成率, 并降低了骨微环境中骨髓基质细胞的成骨分化, 从而导致骨吸收过程增强。 孟唤男等[52]研究发现,骨痛灵、唑来膦酸及二者联合用药通过阻断整合素αvβ3 影响破骨细胞伪足小体的形成, 进而对破骨细胞的骨吸收功能产生一定影响。 韩晓玲等[53]研究指出2-乙酰基毛蕊花糖苷虽然对破骨前体细胞活性没有显著影响, 但能显著抑制巨噬细胞集落刺激因子(macrophagecolony stimulating factor,M -CSF) 和RANKL 共同诱导的抗酒石酸酸性磷酸酶(tartrate resistant acid phosphatase,TRAP)活性增加,进一步表明2-乙酰基毛蕊花糖苷具有抑制破骨细胞形成及其功能的作用。
在畜禽骨代谢的作用机制中,PI3K/Akt 是一条极其重要的信号转导通路, 也是许多信号通路的必经途径。 成纤维细胞生长因子9(fibroblast growth factor 9,FGF9)的体外分化试验表明,FGF9通过共激活MAPK 和PI3K/Akt 信号通路,促进破骨细胞生成并使其成为M-CSF 和RANKL 的潜在共刺激因子, 即可通过促进破骨细胞前体细胞聚集和细胞间融合来促进M-CSF 和RANKL 诱导的破骨细胞分化,从而负向调节骨稳态[54]。 总之,整合素通过PI3K/Akt 信号通路对畜禽骨吸收方面的研究具有重要意义。 PI3K/Akt 是整合素的下游级联反应之一,提示未来应加强整合素通过PI3K/Akt 信号通路对骨吸收发挥的作用研究。
整合素能够触发多种信号通路的激活,PI3K/Akt 被认为是整合素信号通路中的关键信号转导通路,通过激活下游靶向信号转导而发挥作用。 Li等[55]研 究 表 明,血 小 板 因 子4(platelet factor 4,PF4)作用于卵巢摘除模型大鼠后下调了5 个成骨关键调节因子的表达水平, 并抑制了整合素α5-FAK-ERK 通路,使成骨细胞减少,骨密度降低,骨微结构恶化, 提示PF4 可能与抑制骨形成引起的骨质疏松有关。 此外,有报道称,ⅠA 类磷脂酰肌醇3-激酶在整合素αvβ3 下游被激活, ⅠA 类PI3K/Akt 途径调节对破骨细胞骨吸收有至关重要的细胞机制,ⅠA 类PI3K 的特异性阻断可能有助于预防骨质疏松症和类风湿性关节炎等疾病中由活化的破骨细胞引起的过度骨质流失[56]。 综上,PI3K/Akt 信号通路在畜禽骨吸收方面发挥重要作用。
4 小结
近年来,整合素通过PI3K/Akt 信号通路对骨骼健康的影响已被人们广泛报道, 但是整合素通过PI3K/Akt 途径参与疾病的作用机制较复杂,作用范围较广泛。通过回顾以往的研究笔者发现,整合素通过PI3K/Akt 信号通路既可以在骨形成过程中促进成骨细胞发挥重要作用, 也能在骨吸收过程影响破骨相关因子的表达。 随着畜禽养殖规模化水平不断提高,畜禽的生长速度加快,畜禽骨骼健康问题也逐渐凸显。 虽然已有研究证明整合素通过PI3K/Akt 信号通路对畜禽骨代谢发挥关键作用,但具体的调控机制仍然不清楚。 因此,关注整合素如何通过作用于FAK/PI3K/Akt 信号通路在畜禽骨代谢调节中发挥关键作用, 对于明确畜禽骨健康关键的调控机制和筛选靶标药物具有重要意义和价值。
猜你喜欢
中国骨质疏松杂志(2024年2期)2024-03-19
中国骨质疏松杂志(2021年9期)2021-10-08
癌变·畸变·突变(2020年4期)2020-08-05
中国临床医学(2019年3期)2019-01-04
安徽医科大学学报(2016年12期)2017-01-15
中国民族医药杂志(2016年6期)2016-05-09
中国医科大学学报(2015年10期)2015-03-01
癌变·畸变·突变(2015年3期)2015-02-27
福建医科大学学报(2014年6期)2014-03-20
中国医学科学院学报(2012年3期)2012-03-25
